The labrisomids, or scaled blennies, are mostly
small and inconspicuous fishes commonly found on
and around Caribbean reefs. They comprise one of
the largest families of New World reef fishes, with
more than 50 regional species and many more cryptic
species (distinct genetic lineages with subtle morphological
differences) waiting to be described. Labrisomid
genera tend to be speciose and species-level identifications
are generally difficult. Fortunately, almost all
the Atlantic labrisomids belong to just four genera.
The larger and more conspicuous members belong to
Labrisomus,
presently with eleven Caribbean species (including
two recently described species from Brazil but found
widely in the Caribbean), and Malacoctenus,
presently with eight Caribbean species. The two
other genera, Starksia
and Paraclinus,
contain numerous species and are tiny and elusive,
typically well-hidden and rarely encountered or
photographed by divers. The remaining regional labrisomids
are two obscure deep-water genera with a single
species each: Nemaclinus
atelestos, found on deep reef walls, and
Haptoclinus apectolophus, an enigmatic species
found only on the Arrowsmith Banks off the Yucatan
at 1,000 feet deep.
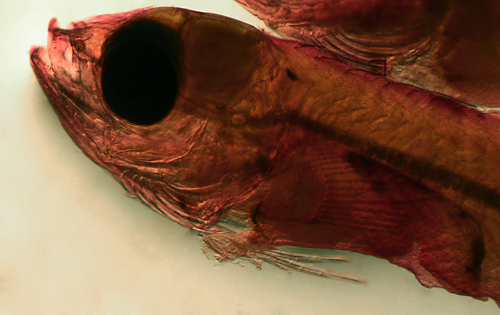
Labrisomid larvae are very common in collections
around Caribbean reefs and are present in more than
90% of my daily larval collections from Panama.
They can be recognized by their long narrow body,
large round eyes, and short pointed snout, with
a long continuous dorsal fin with numerous slender
spines, a long anal fin with just two slender spines,
a very short and narrow caudal peduncle, pelvic
fins thoracic (in front of the pectoral fins) with
only 2 or 3 long strand-like rays (not markedly
curled-up over the body), no obvious head spines,
no silvery peritoneal lining, and light markings.
The markings vary little within the family and typically
comprise a row of melanophores along the anal-fin
base (sometimes also the dorsal-fin base) and a
pattern of spots on top of the head.
These larval characters are shared with the larvae
of the closely-related hole-dwelling blennies of
the family Chaenopsidae,
which are similar in appearance to larval labrisomids
but much less frequent in larval collections. The
chaenopsid blennies are distinguished mainly by
having more dorsal-fin elements (although there
is a small overlap): most chaenopsids have more
than 31 dorsal-fin elements, often with 13 or more
soft rays, while regional labrisomids have 12 or
fewer dorsal-fin soft rays (with a rare 13) and
rarely have more than 32 total dorsal-fin elements.
The Caribbean chaenopsids that can overlap the low
dorsal-fin counts of labrisomids are Coralliozetus
cardonae and the Emblemariopsis
species, along with the rare Emblemaria
vitta. The larvae of these chaenopsids can
be distinguished from similar labrisomid larvae
by their smaller size at stage, fewer markings,
and their mostly differing modes and combinations
of fin-ray counts, as well as fewer procurrent caudal-fin
rays. The taxonomic features generally separating
the two families, i.e. scales on labrisomids and
scales absent on chaenopsids and a set of osteological
characters, are useless for larval stages.
The unusual genus Stathmonotus
is still considered chaenopsid even though their
dorsal fin is made up of all spines and they can
have scales. Some labrisomids of Paraclinus
also have a dorsal fin made up of all spines;
fortunately the larvae of the two genera are easily
distinguished by morphology.
Labrisomid larvae generally resemble those of the
other blennioid families of reef fishes- they can
be distinguished easily from larvae of the combtooth
blennies (family Blenniidae),
which have fewer dorsal-fin spines than soft rays
and blunt snouts at all stages (labrisomids have
twice as many dorsal-fin spines as rays (or more)
and pointed snouts as larvae). Larvae of the blennioid
triplefins (family Tripterygiidae)
have three separate dorsal fins and distinctive
markings and the stargazers (family Dactyloscopidae)
have relatively foreshortened anterior bodies and
curled-up pelvic fins.
Larval labrisomids are superficially similar to
the larvae of gobies
and scarids,
which also often have a similar anal-fin row of
melanophores and are very common in collections
and are about the same size as labrisomid larvae.
However, those larvae notably have many fewer dorsal-fin
spines, short and/or fused pelvic fins, and narrowed
or oddly-shaped eyes, while later-stage labrisomids
have long thread-like pelvic fins and large round
eyes. Larval gerreids (mojarras) are also common
and can be mistaken for labrisomid larvae, however
they have silvery abdominal linings. Larval grunts
(Haemulidae) often have an anal-fin row of melanophores
and can resemble earlier-stage labrisomids, but
they develop a notably short anal fin and characteristic
tail spots.
Species-level
larval identification in Labrisomidae
There is sufficient divergence in appearance
among labrisomid larvae in the region to identify most later-stage
larvae to species and all to the genus level. The exceptions
are those species recently shown by DNA sequencing to be
made up of sets of closely-related species that can be hard
to distinguish, even as adults (i.e. cryptic species). Some
labrisomids, unlike most other reef fishes, have benthic
eggs and short larval lives which promote reproductive isolation
and genetic divergence within the region. As a result, there
can be a proliferation of cryptic species and lineages and
quite complex phylogeography. The larvae and juveniles of
cryptic species would be expected to be almost identical
and are thus pooled into a type for that species complex
in the descriptions below.
Larval identifications are possible
in the labrisomids using a combination of marking
patterns, fin-ray counts, and morphology. Basic body
shape is often useful as a preliminary screen, typically
a gestalt rather than a conscious calculation of proportions,
usually narrowing down the identification to genus.
Melanophore patterns are easily observed and most
useful for separations, but they are rarely species-specific
and show a troublesome propensity to vary. Nevertheless,
in combination with other features, marking patterns
can quickly identify most larvae. Fin-ray counts are
also rarely diagnostic on their own, but can be required
for the identification. Counts can be laborious but
sometimes necessary, especially on ambiguous-appearing
larvae. Specific morphological characters, especially
the relative lengths of fin elements, occasionally
are the deciding factor.
The Diagnosis paragraph under each species
listed in the following sections describes the criteria
that confirm the species designation for a larval
type, usually the fin-ray counts, narrowing the
possibilities to one or a few species and sometimes
a morphological feature to distinguish among the
remainder. Of course, a DNA-sequence match is the
ultimate confirmation. A sequence match has been
used for many of the taxa described below, indicated
by the notation (DNA).
The Analogues section briefly describes
how to distinguish larvae at particular stages from
other similar-appearing species, highlighting which
of the characters- fin-ray counts, melanophore patterns,
morphology, or various combinations thereof- are
most useful for each comparison. In most cases,
especially for late-stage larvae, these features
can rapidly narrow down an ID to the species level.
Procurrent caudal-fin rays
Some diagnostic characters are more easily
visible on larvae than on larger fishes. One of the more
obscure characters that is useful for distinguishing among
the numerous blennioid families and genera is the number
of procurrent rays in the caudal fin. These accessory rays
are defined as the non-segmented rays anterior to the large
segmented caudal-fin rays. The transparency of fish larvae
allows for an easy assessment of the number of procurrent
caudal-fin rays. Adjustment of the transmitted-light angle
highlights bony tissues well. Fortunately, the caudal-fin
rays are usually preserved in otherwise-damaged larvae and
they can provide diagnostic information for identifications
when little else is available.
There is a quite consistent count
of 13 caudal-fin segmented rays in the blennioids,
7 dorsal and 6 ventral, with a variable number of
procurrent rays. Procurrent-ray counts usually vary
within species by one, and usually there is one additional
procurrent ray in the dorsal series than in the ventral
series. Among the labrisomids, Paraclinus
have the fewest, with only 4 or 5, while Starksia
typically have 5 or 6, and Malacoctenus
and Labrisomus
have 6 to 10. Although the numerical differences can
be slight, the procurrent rays look distinctly more
crowded in the latter genera. Other blennioid families
can also be distinguished: chaenopsids
have few, from 3 to 5, while the tripterygiid Enneanectes
have 6 to 8.
Larval melanophores
Labrisomid larvae share, to varying degrees, a
basic set of melanophores that develop progressively
after hatching and are very useful for identifying
larvae at least to genus and often to species. The
timing of the development of each marking can be
somewhat variable within a species, resulting in
a variety of patterns on earlier-stage larvae that
preclude a simple key to species identification.
Late and pre-transitional larvae typically have
their full complement of larval melanophores, making
identifications at these larger sizes somewhat easier.
During transition, however, larval melanophores
begin to disappear and metamorphic melanophore patterns
develop. Metamorphic melanophores consist primarily
of intricate patterns of small surface melanophores,
usually starting on the head and spreading over
the body to form the juvenile markings. This transitional
sequence is also variably timed, promoting a proliferation
of intermediate melanophore complements that can
easily lead to the impression of a greater number
of species than are actually present.
The basic melanophore complement on most labrisomid
larvae comprise the following list, from head to
tail:
note: a number of other melanophores are
present at specific, and often diagnostic, locations
on the larvae of particular species (or groups of
species) and are discussed under the genus or species
descriptions in the next sections.
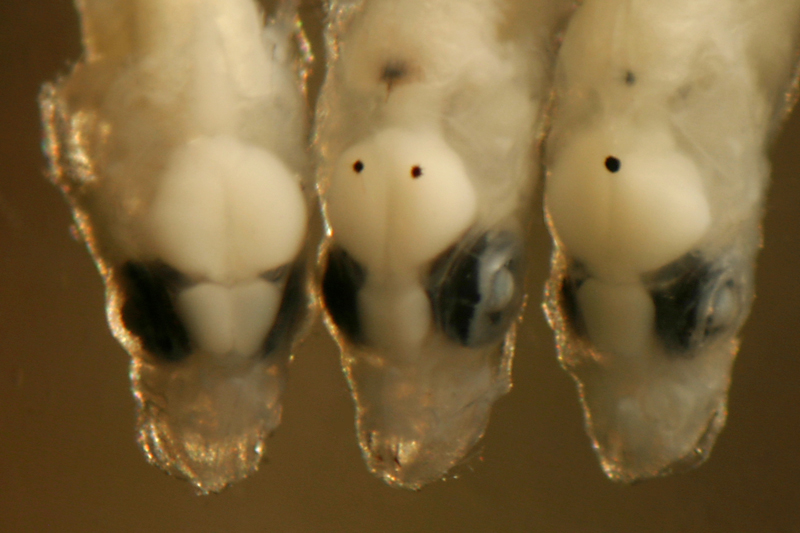
Cranial:
These dorsal melanophores shield the brain and are
near the surface, usually on the meningeal membrane
or over the skull. They can best be labeled by the
quadrant of the brain they overlie, i.e. the forebrain,
the smaller lobes forming a triangle between the eyes,
and the large midbrain (optic) lobes, behind the eyes
(forebrain or midbrain cranial melanophores).
They are present in differing patterns on many of
the larger labrisomid species and are usefully classified
as midline or paired (side-by-side) and uniformly
large or a range of sizes. (left: M. macropus
with paired midbrain cranial melanophores) fc mc
Deep
Nuchal: This large midline melanophore develops
in the early stages at or just after hatching in most
labrisomid genera (deep nuchal melanophore).
It lines the musculature behind the braincase, overlying
the brainstem and the exit of the spinal cord. In
early stages it is near the surface but becomes deeper
and progressively more obscured in larger well-developed
larvae (but still diagnostically important). It is
rarely absent on larvae in those genera after the
very early stages. (left: M. macropus, with
nuchal muscle removed) dn
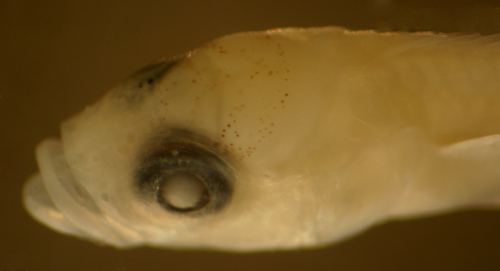
Cheek:
The cheek melanophore lies just under the upper
edge of the preopercle on each side and is characteristic
of some labrisomid genera. The cheek melanophores
develop on later larvae and can be variably absent
on mid-stage larval samples. On well-marked later-stage
larvae of those genera, the cheek melanophores are
virtually always present. ch
Otic:
Larvae of most labrisomid genera have one or two melanophores
on the capsule that surrounds the otoliths, likely
protecting the hair cells of this sensory organ critical
to balance and hearing. These deep internal melanophores
are placed on each side of the lower braincase, just
above the gill cavity, and are visible on early-stage
and more translucent larvae. They are often obscured
on well-developed larvae (otic melanophore).
ot
Isthmus:
The ventral midline forward of the pelvic fins, the
isthmus, can be divided into an anterior portion running
from the cleithral symphysis forward to the hyoids
and a posterior half from the cleithral symphysis
back to the insertion of the pelvic fins (this segment
overlaps the pelvic fin musculature, a triangle with
the apex near the cleithral symphysis). Melanophores
can be located anywhere along the isthmus, but are
usually at the mid-isthmus (around the cleithral symphysis)
or the posterior isthmus, which merges into the pelvic
fin musculature. If the melanophore extends along
the symphysis between the fin insertions, it can be
considered a pelvic-fin melanophore (anterior,
mid-, and posterior isthmus melanophores).
(left: Starksia robertsoni with pinpoint mid-isthmus melanophore)
ai, mi, pi
Pelvic-fin
Base (including Transverse Septum): Melanophores
can be at several locations around the pelvic-fin
base (usually placed well behind the more prominent
isthmus melanophore, as illustrated at left). There
is commonly a deep midline melanophore beneath the
pelvic-fin insertion (deep pelvic melanophore).
On early-stage larvae, this melanophore can be seen
covering the transverse septum, the membrane separating
the pericardial (thorax) and peritoneal (abdominal)
cavities (the pericardioperitoneal membrane), and
it may function to shield the thoracic organs from
sunlight. The deep pelvic melanophore can extend
to the surface at or forward of the pelvic-fin insertion,
or there can be a separate surface melanophore,
often linear, along the pelvic symphysis near the
base of the rays (surface pelvic melanophore).
Less commonly, there can be a melanophore tucked
behind the base of the rays, technically on the
abdomen and sometimes connecting with the transverse
septum (post-pelvic melanophore). As larvae
develop, the deep melanophore can become obscured,
but is still an important diagnostic character.
(left: Paraclinus fasciatus above with mid-isthmus
and deep pelvic melanophores; below Starksia
occidentalis with additional abdominal) dp,
sp, pp
Dorsal
and Posterior Peritoneum: Two or three large
melanophores overlie the abdomen and are usually expanded
to shield the viscera, i.e. along the dorsal aspect
of the swim bladder and extending along the rear wall
of the abdomen to surround the hindgut; both locations
are technically retroperitoneal, just outside the
peritoneal cavity. They are present as a rule on all
labrisomid larvae at all stages, but are not visible
on well-developed larvae (retroperitoneal melanophores).
rp
Anal-fin Base Row:
A row of melanophores along the posterior ventral
midline is present on all labrisomid larvae, typically
one at the base of each anal-fin soft ray and sometimes
at the base of the spines as well. On earlier-stage
larvae, only some of the anal-fin rays have an associated
melanophore and the sequence of development of the
row can be a useful character. Some melanophores
in the anal row can be irregular, either larger
or deeper (often both), and then are an important
diagnostic character. .(anal row melanophores).
(left; Starksia occidentalis with larger
deeper last one)
The total posterior ventral-midline melanophore
count is a useful screening tool for larvae, counting
both anal row and ventral caudal peduncle melanophores.
pvm
Ventral
Caudal Peduncle: The anal-fin row of melanophores
extends past the fin onto the caudal peduncle in many
species, ending at or before the start of the procurrent
caudal-fin rays (non-segmented rays). These melanophores
are often variable in number and placement. Technically,
the melanophore just after the base of the last ray
should be considered part of the anal row if still
touching the ray or pterygiophore (ventral caudal-peduncle
melanophores). vcp
Metamorphic
Melanophores: During the settlement transition,
larvae rapidly develop patterns of tiny surface melanophores
that are usually easily distinguished from the larger
and darker larval melanophores. Larval melanophores,
typically isolated and discrete, extend below the
surface and are often dendritic, while metamorphic
melanophores form intricate patterns, usually species-specific.
The two sets of melanophores can coexist for some
period after settlement, providing a useful link between
larvae and clearly identifiable juveniles. (left:
transitional larva of Malacoctenus triangulatus
with metamorphic melanophores and persistent
cheek and midbrain cranial larval melanophores)
Early-stage larvae
Labrisomids have demersal brooded eggs and hatch
several days after fertilization as well-developed
larvae around 3 mm in length. The early-stage post-flexion
larvae can be recognized by a long, narrow, tapering
body with a small pointed head, medium terminal
mouth, relatively large, mostly rounded eye, a few
small preopercular spines (or none), snout-to-vent
length slightly less than half of body length, a
short caudal peduncle with long dorsal and anal-fin
bases with early-forming posterior elements on the
dorsal fin and inconspicuous slender spines and
rays. Pigmentation follows the genus-level identification
patterns, primarily various head and ventral midline
melanophores. Before larvae develop their full complement
of fin rays and melanophore patterns, species-level
identifications would often require DNA sequencing.
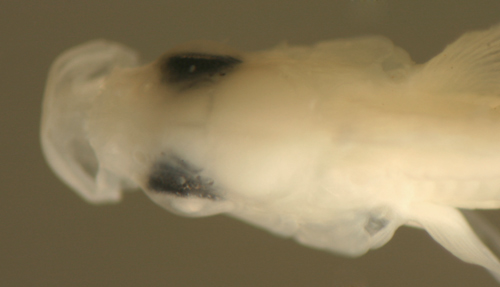
The pigment pattern on the common Malacoctenus
macropus larvae comprises a deep nuchal
melanophore, usually one or two midbrain cranial
melanophores, a cheek melanophore, and the anal
row extending onto the ventral midline of the caudal
peduncle. There are internal otic and retroperitoneal
melanophores.
Malacoctenus macropus
early larvae
5.9 and 7.5 mm SL
-melanophore(s) over
cranium
-one deep behind the braincase
-cheek melanophore
-no anterior ventral midline melanophores
-2-3 small preopercular spines
San Blas, Panama SB86-906
Malacoctenus
vs. Labrisomus
The two more conspicuous labrisomid
genera, Malacoctenus and Labrisomus,
are frequently observed by divers on Caribbean reefs.
The Malacoctenus species are relatively small,
usually an inch or two, and ubiquitous on reefs, while
Labrisomus species are larger and bulkier (some
can reach six inches) and not as common. The two genera
are best distinguished (in collections) from their
tiny relatives, Starksia
and Paraclinus,
by fin-ray counts: Starksia
have fewer dorsal-fin soft rays and pectoral-fin rays
(with a few exceptions) and Paraclinus
have all or all-but-one dorsal-fin elements spinous.
Despite the obvious differences among adult labrisomids
in size and shape, their larvae can appear quite similar
and can be a problem to identify, even to genus.
The larvae of Malacoctenus
and Labrisomus are difficult to separate since
many of their features, including fin-ray counts,
can overlap. In addition, there is little diversity
in larval markings within these genera and many species
have fundamentally similar melanophore patterns. Adding
to the problem, most species show some variation in
the diagnostic patterns of melanophores, especially
the cranial melanophores. Many individuals show less
than the full complement characteristic of the species
and, occasionally, a variant specimen will have a
set of additional markings, notably overlapping the
patterns characteristic of other species. I have confirmed
this with DNA sequencing, although it is uncommon.
DNA sequencing is probably necessary for a firm species
identification in some groups and certainly in many
earlier stages.
Markings:
Most of the variation in appearance among the larvae
of Malacoctenus and Labrisomus is
in the pattern of melanophores on the head and the
presence or absence of melanophores along the fin
bases. Otherwise, much of the larval morphology and
most of the marking patterns are shared among the
many members of these two large genera and diagnostic
differences can be quite subtle. Fortunately, this
problem is mitigated to some degree by the persistence
of larval melanophores through transition, and often
well into the juvenile stage, providing useful links
to establish species identification.
In addition to the basic set of labrisomid melanophores
discussed above, some species, or sets of species,
have distinctive patterns of larval melanophores
that are indispensable for identifications. A very
few melanophore locations are unique to larvae of
one species, but combinations are often diagnostic
for species. An interesting observation is that
melanophores that are characteristic of one species
may occasionally (or rarely) be found on other related
species to a lesser degree or at a later point in
transition. Because of these overlaps, not every
larva can be easily assigned to species on the basis
of a diagnostic larval melanophore pattern; several
characters sometimes need to be weighed in the decision
(note that metamorphic melanophore patterns
are very specific and diagnostic). No single marking
identifies all Malacoctenus or Labrisomus,
but some are found only in one genus (but not in
all of the species), such as the pair of melanophores
behind the tip of the upper jaw on some Labrisomus.
Morphology:
Most of the adult characters that separate Malacoctenus
and Labrisomus do not apply to larvae.
Adults are typically separated by the shape of the
snout: pointed in Malacoctenus and blunted
in most Labrisomus (except for very pointed
in L. nigricinctus). This feature does not
apply to the larval stages (or even small juveniles)
which have pointed snouts in both genera. Most Labrisomus
species develop characteristically thick bulky
heads as juveniles and adults, but this too does not
apply to the larval stages. Another basic difference
in adult fishes is that the rear edge of the maxilla
is sheathed in Malacoctenus and exposed in
Labrisomus, but this is difficult to assess
on larvae.
The mouth is typically small in both juvenile and
adult Malacoctenus (and L. nigricinctus)
and the maxilla extends back only to the level of
the anterior orbit vs. large and extending back
past the level of the pupil in most Labrisomus.
This mouth-size character is the best character
for distinguishing juveniles but does not generally
apply in larvae, and, when it does, can be quite
subtle, although sometimes useful. For example,
in the photograph below, the maxilla is distinctly
longer in the 15 mm Labrisomus
guppyi larva on the left vs. the same-sized
Malacoctenus
triangulatus on the right (note eye diameters
are identical).
The dorsal-fin profile typically differs between
genera (and often species). A loose but useful rule
for juveniles and adults is that most Malacoctenus
have very short penultimate and third-to-last dorsal-fin
spines, less than a third the length the longest
dorsal-fin soft ray, while many Labrisomus
have the shortest dorsal-fin spine about half or
more the length of the longest dorsal-fin soft ray.
There are certainly exceptions: L.
nigricinctus, L.
albigenys, and, to a degree, the 19-spined
Labrisomus species have short posterior
dorsal-fin spines, and M.
macropus and M.
boehlkei have relatively long posterior
spines. The dorsal-fin profiles of larvae do not
always match those of juveniles or adults, but they
can be helpful, especially for late larvae approaching
transition. In general, most Labrisomus
larvae have relatively low soft dorsal fins while
many Malacoctenus larvae have high soft dorsal
fins and short posterior dorsal-fin spines (see
transitional larvae below: Labrisomus
haitiensis top vs. Malacoctenus
versicolor bottom).
Transition:
Larvae undergo a rapid and profound transition during
the night of settlement as they prepare for life on
the reef after living as transparent pelagic larvae
in the open ocean. The changes include a new set of
markings, longer fins, cirri on the head, and changes
in body shape. The genus-level larval differences
in mouth size and dorsal-fin profile become more developed.
Malacoctenus larvae (right and below) develop
distinctly longer cirri early in transition, with
the orbital and nuchal cirri often more than a lens
diameter in length, while Labrisomus larvae
have short and stubby cirri through the transition,
although later, as juveniles, their cirri become long
and prominent.
In contrast to the similarity in
larval markings, the patterns of fine surface melanophores
that develop at metamorphosis are often complex and
distinctive and thus transitional larvae and recruits
can be assigned to species relatively easily. These
metamorphic melanophores are usually easy to differentiate
from larval melanophores by their much smaller size,
their lighter appearance (often appearing brown vs.
black), and their occurrence in dense patches or in
a dense uniform speckling (occasionally in long strands
or lines). Technically, they can be defined as tiny
melanophores (about 10-25 microns in diameter, or
about 30 into the lens diameter) with multiple similar-sized
melanophores within two diameters of each spot. The
M.
triangulatus larvae illustrated (upper left)
has both arrays. Occasionally, some transitional larvae
can develop a few additional melanophores that look
just like small larval melanophores, i.e. darker and
2 or 3 times the size of metamorphic melanophores
(shown on M.
macropus at lower left). I refer to these
troublesome melanophores as "pseudo-larval"
melanophores. In some species, they occur in clusters
around fading large larval melanophores, resembling
a "fragmentation" of the larval melanophores.
An interesting pattern often observed is the tendency
of metamorphic melanophores to leave a clear halo
around larval (and pseudo-larval) melanophores.
Another general, although less
reliable, difference between the two genera is the
hunched-over appearance that develops in many transitional
Labrisomus larvae (e.g. L.
bucciferus at right and L.
haitiensis full-body photo above), where the
head is generally lower than the body: i.e. the tip
of the jaw (and often the center of the orbit) is
below the lateral midline of the body. There is also
an overall size difference between late larvae, with
Malacoctenus generally settling between 10
and 15 mm SL and most Labrisomus settling between
15 and 20 mm SL. There is, however, some overlap between
the larger-settling Malacoctenus (e.g. M.
triangulatus and M.
versicolor) and the Labrisomus species.
The morphology of the
two genera diverges rapidly after transition and juveniles
can be easily distinguished. Most juvenile Labrisomus
have thick bulky heads and large mouths with the maxilla
extending past the midline of the orbit and juvenile
Malacoctenus are slim and have a small mouth.
In addition, Labrisomus juveniles have dark
or mottled camouflage and are usually well-hidden
in their habitats while Malacoctenus are usually
out in the open and have distinctive colors and patterns.
Malacoctenus and Labrisomus
species list
Fin-ray
counts: Although there is extensive overlap,
fin-ray counts are very useful in this family since
the modes within species are relatively strong, often
two-thirds or more of the specimens, and the ranges
of counts are well-documented. Note, however, that
recent evidence indicates that local cryptic species
or populations can have slightly variant modes. The
species are listed here in order of increasing fin-ray
counts (the range, with the known combinations in
parentheses).
Counts are mostly from Springer's
classic monograph:
Springer, V. G.: Systematics and zoogeography of
the clinid fishes of the subtribe Labrisomini Hubbs.
Pubs. Inst. Mar. Sci. Univ. Texas 5:417–492
(1959), often cited as (1958).
Malacoctenus
M.
versicolor: mode D-XVIII,12 and A-II,18-19
P-14 (rarely 11 dorsal-fin soft rays; P-13-14)
M.
delalandii: mode D-XX,10 and A-II,19 P-14
(dorsal 19/9-11, 20/9-11, 21/9; A-II,17-20; P-13-15)
M.
gilli: mode D-XX,10 and A-II,19 P-14 (dorsal
18/10, 19/10-11, 20/9-11, 21/9-10; A-II,17-21; P-13-16)
M.
aurolineatus: mode D-XIX-XX,11 and A-II,19-20
P-14 (dorsal 18/10-11, 19/10-12, 20/10-12, 21/10-11;
A-II,17-21; P-13-15)
M.
triangulatus: mode D-XX,12 and A-II,21 P-14
(dorsal 19/12-13, 20/11-13, 21/11; A-II,20-22; P-13-15)
M.
erdmani: D-XXI,9 and A-II,18-19 P-16 (dorsal
20/9-10, 21/8-10, 22/8-9; A-II,17-20; P-15-17)
M.
macropus: D-XXI,9-11 or XXII,8-10 or XXIII,9-10
and A-II,20-21 (range 18-22) P-15 (range 14-16)
M.
boehlkei: D-XX,13 or XXI,11-12 or XXII,11
and A-II,22 (range 20-23) P-15
note: Some Starksia
species can barely overlap the lower range of fin-ray
counts for those few Malacoctenus species
that can have 8 or 9 dorsal-fin soft rays and 14
or fewer pectoral-fin rays.
Labrisomus
18 dorsal-fin spine group
L.
albigenys: mode D-XVIII,11 A-II,18 P-13 (holotype
recorded as D-XVIII,10 A-II,18, not repeated)
note: some chaenopsid
blennies overlap the fin-ray counts of many of the
labrisomids listed above: i.e. the genus Emblemariopsis
and Coralliozetus
cardonae (D-XVIII,12 A-II-20 (dorsal 17-19/10-13;
A-II,18-24; pectoral 11-13) and (rarely) Emblemaria vitta (D-XVIII-IXX,13-14 and
A-II,19-20).
Malacoctenus
presented in order of increasing dorsal-fin elements
Malacoctenus versicolor
Diagnosis: The modal
fin-ray count of D-XVIII,12 A-II,18 and P-14 indicates
Malacoctenus versicolor and several Labrisomus
species, including L.
nuchipinnis, L.
conditus, and L.
cricota. M. versicolor can be distinguished
from the latter species by the dorsal-fin profile,
with the first spine always the longest (vs. shorter
than the others in L.
nuchipinnis and L.
conditus) and the third-to-last dorsal-fin
spine short (three times into the longest soft ray)
vs. half or more in the three Labrisomus,
as well as a smaller mouth (maxilla not past the
midpoint of the eye). The 19-spined
Labrisomus species can sometimes overlap
the fin-ray count, but they also have distinctly
larger mouths (maxilla past the midpoint of the
eye). The fin-ray count also (barely) overlaps with
the chaenopsid blenny Coralliozetus
cardonae, which are much smaller at all
stages and have only 3-4 procurrent caudal fin rays
(vs. 6-8). (DNA)
Ecology:
The barfin blenny is a small blenny found in shallow
coral and rocky areas and mixed habitats with complex
structure and uncommonly observed or photographed
by divers. Indeed, photographs on the web and in
guidebooks are often the barred phase of M.
macropus, including those in Humann's book
and on the Smithsonian larval-fish website. Genuine
barfin blenny photographs include those from St.
Vincent by Keri Wilk (ReefNet) and from Eleuthera,
Bahamas by Louis Johnson. The species is found mainly
in Florida, the Bahamas, and the northern Caribbean
islands; it is not recorded from the Gulf of Mexico
or Bermuda and appears to be replaced by M.
delalandii to the south of Belize and the
lower Antilles, i.e. in Panama to Venezuela and
mainland Brazil. Barfin blennies are infrequent
in collections and are found in numbers only in
rare localities, especially in the Bahamas. Their
larvae are rare in collections.
Description: Pre-transitional larvae:Body long,
moderately narrow, and thin with a medium round
eye, pointed snout, and small terminal mouth. Long
continuous dorsal and anal fins with a very short
and narrow caudal peduncle. Pectoral fins long,
reaching past the vent, and pelvic fins long and
thread-like. On the head there are several large
melanophores along with several smaller melanophores,
overlying all quadrants of the fore- and midbrain,
usually more than 5 per side. There is a cheek melanophore
on each side. There are no melanophores along the
base of the dorsal fin, but there are several melanophores
at the base of the upper and lower caudal-fin segmented
rays. Along the ventral midline there is a melanophore
at the isthmus and deep at the pelvic-fin base.
Along the anal fin there is a melanophore at the
base of each anal-fin soft ray, closely followed
by two or three along the ventral midline of the
caudal peduncle. Internal melanophores comprise
only the basic complement: the nuchal midline, otic
capsule, and overlying the abdominal organs.
Transitional stage: M. versicolor
larvae in transition develop lines and well-outlined
patches of small surface melanophores over the head,
notably with two thin lines extending down and slanted
forward from the orbital rim at 6 o'clock. The metamorphic
melanophores on the body form complex shapes and
rings, some of which connect into narrow bars that
extend uninterrupted over the dorsal fin to the
edges of the membranes. Multiple long cirri form
on the nape, over the eye, and over the nasal tube.
Juveniles: M. versicolor juveniles
have relatively narrow dark bars on the body that
extend across the dorsal-fin membranes, notably
not widening as they reach the edge of the fin.
The bar under the last dorsal-fin spine is characteristically
narrow and unbranched. The first three dorsal-fin
spines are distinctly longer than all subsequent
spines.
Analogues:
Several other Malacoctenus species can have
larvae with the head speckled with 10 or more spots.
M.
triangulatus larvae usually have many more.
The two species are similar in size and morphology,
but can be separated by fin-ray counts: 18 dorsal-fin
spines and 18-19 anal-fin soft rays in M. versicolor
vs. 20 dorsal-fin spines and 20-21 anal-fin soft
rays in M.
triangulatus. In addition, M. versicolor
larvae have an obvious third pelvic-fin ray about
two-thirds the length of the second vs. less than
half the second and often inconspicuous in M.
triangulatus and a row of melanophores along
the caudal-fin base vs. none or a single melanophore
(at the base of the largest dorsal procurrent ray)
in typical M.
triangulatus larvae. M.
gilli larvae can occasionally have similar
numbers of head spots, but most have 20 dorsal-fin
spines and they are smaller and lightly marked,
with no melanophores along the caudal-fin base.
M.
boehlkei larvae also have numerous head
spots, but have 33 dorsal-fin elements.
Since the markings and size of M. versicolor
larvae can be intermediate between typical Malacoctenus
and Labrisomus, separation from Labrisomus
larvae can be problematic. Several of the Labrisomus
species that share fin-ray counts with M. versicolor
are slimmer forms with smaller mouths that
can share morphology with Malacoctenus: i.e.
L.
nuchipinnis, L.
conditus, L.
cricota, as well as the two small species,
L.
nigricinctus and L.
albigenys; the latter two with fewer head
melanophores and much shorter first dorsal-fin spines
than M. versicolor. Larvae of L.
nuchipinnis, L.
conditus, and L.
cricota differ primarily in having melanophores
along the bases of the spinous and soft dorsal fins
and a prominent U, V, or O-shape arrangement of
large melanophores over the head. The 19-spined
Labrisomus species can sometimes overlap
the 18 dorsal-fin spine count, but they typically
have melanophores along the dorsal-fin base and
behind the tip of the upper jaw, as well as a different
dorsal-fin profile as larvae, with shorter first
spines and relatively longer posterior spines. L.
haitiensis larvae share all of these differences
(except the shorter first dorsal-fin spine) and
also have higher fin-ray counts.
Transitional M. versicolor larvae are distinguished
by fin-ray counts, persistent larval melanophores,
and their metamorphic melanophore pattern, especially
the narrow bars on the body vs. wide inverted triangles
in transitional M.
triangulatus and generally wider bars or
triangles, covering four or more dorsal-fin-spine
bases, in the other barred Malacoctenus species.
Some transitional M.
macropus have a pattern of of ovals, but
not organizing into long bars. Most transitional
Labrisomus larvae have uniform metamorphic
melanophore patterns without the sharply-outlined
shapes of M. versicolor. The exception is
L.
haitiensis, whose larvae develop a similar
pattern of well-delineated shapes, however their
shapes do not connect and form long narrow bars
(and they have higher fin-ray counts, a short third
pelvic-fin ray, and a larger mouth).
Juvenile M. versicolor are distinguished
by their prominent narrow dark bars on the body
which extend onto the dorsal fin membranes, but
several other related species can share this character.
An uncommon barred
variant of juvenile M.
macropus has the bars, but are distinguished
by having long single cirri over the nape, eye,
and nasal tube (vs. multiple in all congeners) and
the bars under the last spines and first rays merge
to form a Y or V-shape and the bars are often limited
to the dorsal aspect. M.
delalandii can appear very similar, sharing
the bar pattern extending onto the dorsal fin, but
their dark bars distinctly widen, meeting (or almost
meeting) at the edges of the fin. They also have
relatively shorter first dorsal-fin spines (the
mid-fin spines are longer than the first) and one
or two more dorsal-fin spines. Among the Labrisomus,
L.
cricota is the most similar to M. versicolor,
with similar markings and also with a long first
dorsal-fin spine, but the mouth is larger (maxilla
past the midpoint of the eye), the second bar on
the dorsal fin slants down and forward to the operculum,
and the last bar on the spinous segment of the dorsal
fin covers only spines (vs. the base of rays as
well). L.
conditus and L.
nuchipinnis have short first dorsal-fin
spines and the aforementioned bar differences. L.
nigricinctus juveniles have prominent bars,
but have an obvious large opercular ocellus. Other
Labrisomus species can have similar
narrow bars on the body extending onto the fins,
but are separated by their blunt snouts and large
mouths.
Malacoctenus versicolor
transitional larva
15.6 mm SL, DNA-confirmed
ID
St. Thomas, USVI/J. Lamkin,
A. Shiroza
Malacoctenus versicolor
juvenile
Eleuthera, Bahamas
courtesy Louis Johnson
Malacoctenus versicolor
juvenile
Eleuthera, Bahamas
courtesy Louis Johnson
Malacoctenus delalandii
Diagnosis: The modal
fin-ray count of D-XX,10 A-II,19 and P-14 indicates
Malacoctenus delalandii or M.
gilli. Many M.
aurolineatus share this fin-ray count, but
their mode is 11 soft dorsal-fin rays. Some M.
erdmani share the median-fin ray count but
have a mode of 16 pectoral-fin rays. L.
kalisherae, L.
bucciferus, and L.
haitiensis can also overlap the fin-ray
count. The species is often spelled as Malacoctenus
delalandei. (DNA)
Ecology:
The Brazilian blenny is found mostly in the southern
Caribbean, where it seems to replace M.
versicolor. The species has been collected
from the inshore reefs of Belize south along the
Central American coastline and across to NE Venezuela,
where it is the most abundant species of the genus.
Their range extends south to mainland Brazil, but
not to the offshore islands of Noronha and Rocas.
They are small blennies found on inshore shallow
reef areas with complex structure; within most of
their Caribbean range they are not commonly observed
by divers. Their larvae are unknown or unrecognized
in collections.
Description: (larvae
unknown)
Juveniles:M. delalandii
juveniles have dark bars on the body that extend
uninterrupted over the full-width of the dorsal
fin. The dark bars on the fin membranes distinctively
expand outwards, often meeting at the fin edge.
There is a dark spot on the lower operculum, often
outlined and elongated, but not an obvious round
ocellus.
Analogues:
Juveniles of M.
versicolor can appear quite similar, but
their first dorsal-fin spine is notably longer than
the rest (vs. M. delalandii with the first
spine about equal or shorter than the mid-fin spines),
the bars that extend onto the fins do not expand
to meet at the edges, the bar under the last dorsal-fin
spines is narrow and separate from the next dark
patch forward (vs. widening anteriorly as it spreads
onto the fin in M. delalandii), and the dorsal-fin
spine count is lower. An uncommon barred
variant of juvenile M.
macropus can also have bars on the body
extending uninterrupted over the dorsal fin, but
they also do not expand to meet at the edges, their
fin-ray counts are different, and they have long
single cirri (vs. multiple in M. delalandii
and M.
versicolor). Labrisomus
nigricinctus can share the barred pattern,
but they have an obvious round opercular ocellus.
Malacoctenus
15.4 mm SL
San Blas, Panama, SB81-074
Malacoctenus gilli
Diagnosis: The modal
fin-ray count of D-XX,10 A-II,19 and P-14 indicates
Malacoctenus gilli and M.
delalandii. Some M.
aurolineatus can share this fin-ray count,
but most have 11 soft dorsal-fin rays. Some M.
erdmani share the median-fin ray count but
have a mode of 16 pectoral-fin rays. L.
kalisherae, L.
bucciferus, and L.
haitiensis can also overlap the fin-ray
count. (DNA)
Ecology:
The dusky blenny is a somewhat common small blenny
found primarily in shallow mixed habitats in bays.
They have a curious, mostly island, distribution,
found in the east from Bahamas and the Antilles
down to offshore Venezuela and then in the west
in Yucatan, Belize, and some offshore islands, but
apparently not in Florida, Panama and the SW Caribbean
or NE Venezuela. Their larvae are occasional in
collections.
Description: Pre-transitional larvae: Body long,
narrow, and thin with a large round eye, pointed
snout, and relatively small terminal mouth. Long
continuous dorsal and anal fins with a short and
narrow caudal peduncle. Pectoral fins long, reaching
past the vent, and pelvic fins long and thread-like.
The complement of melanophores on the top of the
head is quite variable, with one to several larger
melanophores overlying the midbrain lobes and sometimes
additional smaller melanophores over the forebrain
lobes (may be pseudo-larval: developing during transition,
but look like larval melanophores). There is a cheek
melanophore on each side. There are no melanophores
along the base of the dorsal or caudal fins. Along
the ventral midline there is notably no isthmus
melanophore, but there is a deep melanophore behind
the pelvic-fin base. Along the anal fin there is
a melanophore at the base of each anal-fin soft
ray (often not the last), followed by one or two
melanophores along the ventral midline of the caudal
peduncle. Internal melanophores comprise only the
basic complement: the nuchal midline, otic capsule,
and overlying the abdominal organs.
Transitional stage: M. gilli
larvae in transition first develop patches of small
surface melanophores over the head, including a
long bar down from the orbital rim at 5 o'clock
and a broad eye-stripe from the orbital rim to the
mid-maxilla. There are often several small pseudo-larval
melanophores over the cranium in addition to the
fine surface array. The fine melanophores on the
pectoral-fin-base develop first as two small patches
along the lower margin of the fleshy pectoral-fin
base. Fine metamorphic melanophores later extend
onto the body, forming complex patches and often
arrays of small dark spots and a large dark spot
forms on the first two dorsal-fin spine membranes.
Multiple long cirri form on the nape, over the eye,
and over the nasal tube.
Juveniles: M. gilli juveniles
have very distinctive markings, including a large
black spot over the first two dorsal-fin spine membranes
and an ocellated spot with a blue center over the
rear spinous dorsal fin which extends well onto
the body.
Analogues:
The variable occurrence of one to several large
spots over the rear cranium and/or additional smaller
spots forward overlaps the head pattern found on
most other congeners. Reduced-complement M. gilli
are the only Malacoctenus larvae that share
the bare-forebrain-lobes pattern characteristic
of M.
macropus and M.
erdmani. Fortunately, the anterior ventral
midline series differs: M. gilli larvae have
a deep pelvic-fin base melanophore but no isthmus
melanophore while the remaining Malacoctenus
have both (most species) or neither (M.
macropus and M.
erdmani). M.
versicolor and M.
triangulatus usually have more numerous
head melanophores, often 10+ per side. M.
versicolor and M.
aurolineatus larvae also differ by having
melanophores along the caudal-fin base. The D-XX,10
combination occurs in about two-thirds of M.
gilli individuals, but is uncommon among congeners
other than M.
delalandii. M.
boehlkei larvae have multiple head spots
in alll quadrants, but have 33 dorsal-fin elements.
Early transitional M. gilli larvae are distinguished
by fin-ray counts, persistent larval melanophores,
and their metamorphic melanophore pattern, i.e.
the combination of the 5 o'clock bar of melanophores,
an eye stripe to the mid-maxilla, and two patches
of melanophores along the lower edge of the pectoral-fin
base. M.
triangulatus have a quite similar marking
pattern at transition, but have a single central
patch along the lower rim of the pectoral-fin base;
M.
macropus are also similar but have a stripe
across the lower pectoral-fin base (and single cirri).
Once recruits develop the large dark spot at the
front of the dorsal fin and the characteristic ocellus
on and below the rear spinous dorsal fin, they are
easily distinguished from all other labrisomids.
Ecology:
The goldline blenny is a common small blenny found
primarily in exposed shallow eroded limestone habitats
and mixed coral substrates. They can be found in
Florida, the Bahamas, at the mouth of the Gulf of
Mexico and all of the Caribbean Sea, except NE Venezuela.
Their larvae are occasional in collections.
Description: Pre-transitional larvae: Body long,
moderately narrow, and thin with a large round eye,
pointed snout, and small terminal mouth. Long continuous
dorsal and anal fins with a short and narrow caudal
peduncle. Pectoral fins long, reaching past the
vent, and pelvic fins long and thread-like. On the
head there is a large midline melanophore overlying
the midbrain lobes and usually another single large
midline melanophore over the forebrain lobes; less
often, the anterior melanophores can be smaller,
off-center, or paired. Occasionally there are one
or a few additional small associated melanophores,
although the total number of larval melanophores
on top of the head rarely exceeds 5. There is a
cheek melanophore on each side. Along the dorsal
fin there is a melanophore at the base of some or
all of the soft rays (occasionally also at the base
of one or two of the last dorsal-fin spines). There
are small melanophores in a thin line along the
base of the caudal-fin rays, often both procurrent
and segmented. Along the ventral midline there are
melanophores at the anterior isthmus as well as
at the mid-isthmus at the cleithral symphysis, in
addition to deep just behind the pelvic-fin base.
Along the anal fin there is a melanophore at the
base of each anal-fin soft ray (often not the last),
closely followed by a series (up to 4) along the
ventral midline of the caudal peduncle up to the
procurrent caudal-fin rays. A row of internal melanophores
overlies the vertebral column, spaced every 2-4
vertebrae, extending to the caudal peduncle, where
the melanophores can overlie each vertebral body
(often inconspicuous in stout-bodied larvae). Additional
internal melanophores include those at the nuchal
midline, otic capsule, and overlying the abdominal
organs.
Transitional stage: M. aurolineatus
larvae in transition develop patches of small
surface melanophores over the head, including a
short bar slanting forward and down from the orbital
rim at 6 o'clock (and no obvious eye-stripe from
the orbital rim to the mid-maxilla). In addition,
a distinctive long, thin, and straight vertical
bar forms on the pectoral-fin base. Fine metamorphic
melanophores later extend onto the body, forming
complex patches resembling inverted triangles. Multiple
long cirri form on the nape, over the eye, and over
the nasal tube.
Juveniles: M. aurolineatus
juveniles have an H-pattern of two broad connecting
dark bars on the anterior body and a lighter rear
body. They notably have no large dark spot at the
front of the dorsal fin, on the operculum, or on
other fins. Two prominent long dark vertical lines
on the pectoral-fin base are diagnostic in well-marked
individuals.
Analogues: M. aurolineatus larvae can be distinguished
from other labrisomid larvae by having fewer than
5 melanophores on the top of the head combined with
melanophores along the soft dorsal-fin base and
caudal-fin base. The other Malacoctenus species
with fewer than five melanophores on top of the
head, M.
macropus, M.
erdmani, and sometimes M.
gilli, have no melanophores along the dorsal
or caudal-fin bases. The most common configuration
of two large midline melanophores spaced well apart
on top of the head and two midline melanophores
along the isthmus is not shared by other labrisomid
larvae. Some Labrisomus larvae also have
melanophores along the soft dorsal-fin base, but
they always have more speckled heads than M.
aurolineatus. L.
albigenys and L.
nigricinctus larvae also have few head melanophores
as well as isthmus and pelvic-fin base melanophores
and are similar in size and shape, but the former
have a distinctive enlarged anal-fin base melanophore,
a side-by-side pair of melanophores over the rear
cranium, and no caudal peduncle ventral midline
spots, and both have no caudal-fin base or soft
dorsal-fin base melanophores (and usually lower
fin-ray counts).
Early transitional M. aurolineatus larvae
are distinguished by fin-ray counts, persistent
larval melanophores, and their metamorphic melanophore
pattern, i.e. the combination of the 6 o'clock bar
slanting forward, the absence of an eye stripe to
the mid-maxilla, and, most distinctive, a long,
narrow, and straight vertical line on the pectoral-fin
base that is not shared by other transitional labrisomid
larvae.
Juveniles later develop a second and separate long
vertical line on the pectoral-fin base (similar
lines in M.
gilli are linked with a crossbar in an H-pattern
or form a Y in M.
triangulatus), broad inverted-triangle bars
connected in an H-shape on the anterior body, and,
notably, the absence of large spots or ocelli on
the fins.
Malacoctenus aurolineatus
larva
12.3 mm SL
San Blas, Panama, SB86-429
Malacoctenus aurolineatus
trans larva
12.7 mm SL
San Blas, Panama, SB86-408
Malacoctenus aurolineatus
trans larva
14.3 mm SL, DNA-confirmed
ID
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Malacoctenus aurolineatus
trans larva
14.3 mm SL, DNA-confirmed
ID
cutaway shows vertebral
melanophore row
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Malacoctenus aurolineatus
trans larva
14.3 mm SL, DNA-confirmed
ID
diagnostic vertical line
on pectoral-fin base
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Malacoctenus aurolineatus
trans larva
14.3 mm SL, DNA-confirmed
ID
cutaway shows vertebral
melanophore row
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Malacoctenus aurolineatus
new recruits
14.4, 12.2, 12.2 mm SL
DNA-confirmed ID
St. Thomas, USVI, ST953
Colon, Panama, N7529b
Malacoctenus aurolineatus
new recruit
14.4 mm SL, DNA-confirmed
ID
St. Thomas, USVI, ST953
Malacoctenus triangulatus
Diagnosis: The modal
fin-ray count of D-XX,12 A-II,20-21 and P-14-15
indicates Malacoctenus triangulatus and falls
within the range for M.
aurolineatus, L.
bucciferus, and L.
haitiensis. (DNA)
Ecology:
The saddled blenny is the most commonly observed
labrisomid on coral reefs in the region (M.
macropus are generally more common but they
are less conspicuous and less associated with coral
habitats). Saddled blennies have one of the widest
of distributions for regional reef fishes; they
are found in Bermuda, Florida, the Gulf of Mexico,
the Bahamas, all of the Caribbean Sea including
NE Venezuela, as well as in Brazil and on many of
its offshore islands. Their larvae are common in
collections.
Description: Pre-transitional larvae: Body long,
moderately narrow, and thin with a large round eye,
pointed snout, and small terminal mouth. Long continuous
dorsal and anal fins with a short and narrow caudal
peduncle. Pectoral fins long, reaching past the
vent, and pelvic fins long and thread-like. The
full complement of melanophores on the top of the
head comprises a scattering of large spots grading
down to small, even tiny, spots (typically 10 or
more per side, often many more), sometimes with
the large spots arranged in two inward-facing crescents
in a U- or O-shape. Occasional larvae have incomplete
complements, sometimes just one or two large melanophores
with one or a few smallerl spots. There is a cheek
melanophore on each side. Typical lightly-marked
larvae have no melanophores along the dorsal-fin
base. Many lightly-marked larvae do have a characteristic
single melanophore near the base of the largest
dorsal procurrent caudal-fin ray, but no additional
caudal-fin base melanophores. (A "dorsal-row"
variant with melanophores at the base of some or
all of the soft dorsal-fin rays and even along the
base of some dorsal-fin spines occurs among some
early transitional larvae. These larvae can also
have melanophores along the base of the caudal-fin
rays. It is unclear whether these larvae represent
a regional variant in the Antilles or are only a
variation during early transition and appear the
same as lightly-marked larvae at earlier stages.)
Along the ventral midline there is a melanophore
at the mid-isthmus (rarely an additional one on
the anterior isthmus) and one deep behind the pelvic-fin
base. Along the anal fin there is a melanophore
at the base of each anal-fin soft ray (sometimes
not the last), followed in some larvae by one to
three spots along the ventral midline of the caudal
peduncle (many lightly marked larvae have none).
A row of internal melanophores overlies the vertebral
column, one per vertebra, along the mid- and rear
body, not continuing onto the caudal peduncle. Additional
internal melanophores include those at the nuchal
midline, otic capsule, and overlying the abdominal
organs.
Transitional stage: M. triangulatus
larvae in transition develop patches of small surface
melanophores over the head, including a short bar
down from the orbital rim at 5:30 o'clock and a
broad eye-stripe from the orbital rim to the mid-maxilla.
In addition, the melanophores along the lower edge
of the fleshy pectoral-fin base form a single patch
midway from the body to the fin-ray insertion, which
later develops into a Y-shaped bar. Fine metamorphic
melanophores extend onto the body forming complex
patches, mostly on the upper body, roughly in the
shape of wide inverted triangles. Multiple long
cirri form on the nape, over the eye, and over the
nasal tube.
Juveniles:M. triangulatus
juveniles develop a large dark spot at the base
of the second and third dorsal-fin spines that extends
onto the body, followed by a row of five wide inverted
triangles extending to the tail.
Analogues:
In general, M. triangulatus larvae can be
distinguished from most other Malacoctenus
larvae by having more than 10 melanophores per side
on the top of the head, a pattern shared only by
some larvae of M.
versicolor. The two species are also very
similar in size and morphology, but fortunately
can be reliably separated by fin-ray counts: 18
dorsal-fin spines and 18-19 anal-fin soft rays in
M.
versicolor vs. 20 dorsal-fin spines and
20-21 anal-fin soft rays in M. triangulatus.
In addition, M.
versicolor larvae have an obvious third
pelvic-fin ray about two-thirds the length of the
second vs. less than half the second and often inconspicuous
in M. triangulatus and a row of melanophores
along the caudal-fin base vs. at most a single melanophore
(at the base of the largest dorsal procurrent ray)
in M. triangulatus larvae (with rare exceptions).
Larval M.
boehlkei also have a short third pelvic-fin
ray, a feature shared only by M. triangulatus
and L.
haitiensis, and can have multiple spots
over each quadrant of the cranium; they are best
distinguished from M. triangulatus by having
relatively long posterior dorsal-fin spines and
usually more fin rays (a rare specimen of M.
triangulatus could have the same count).
Since the markings, morphology, and size of M.
triangulatus larvae can be intermediate between
typical Malacoctenus and Labrisomus,
separation from Labrisomus larvae can be
problematic. This is especially true for those variants
with melanophores along the dorsal and caudal-fin
bases and/or a reduced complement of head melanophores,
thus resembling the pattern for many Labrisomus
larvae. The fin-ray count is helpful, with most
M. triangulatus having a high fin-ray count
of D-XX,12 and A-II,21, fortunately non-overlapping
with several of the slimmer Labrisomus that
can share morphology with Malacoctenus (L.
nuchipinnis, L.
conditus, L.
cricota, as well as the two small species,
L.
nigricinctus and L.
albigenys; the latter two with many fewer
head melanophores than M. triangulatus).
The most similar Labrisomus larva is L.
haitiensis, which can share both the high
fin-ray counts and the inconspicuous short third
pelvic-fin ray and its head melanophore pattern
of a U-shape of six large spots can overlap with
reduced-complement M. triangulatus larvae
(and the occasional L.
haitiensis larva have a few additional small
spots). L.
haitiensis larvae, however, always have
the row of melanophores along the dorsal-fin base
and a pair of prominent melanophores near the tip
of the upper jaw (the former rare in M. triangulatus)
and have a different dorsal-fin outline, with longer
posterior spines. Some of the remaining 19 and 20-spined
Labrisomus species can overlap in fin-ray
counts with M. triangulatus, but those larvae
have an obvious third pelvic-fin ray which is inconspicuous
in M. triangulatus and often uniformly sized
head melanophores (vs. graded tiny to large).
Transitional M. triangulatus larvae are
distinguished by fin-ray counts, persistent larval
melanophores, and their metamorphic melanophore
pattern, i.e. the combination of the 5:30 o'clock
bar, an eye stripe to the mid-maxilla, and a single
patch of melanophores (later developing into a Y-shape)
along the lower edge of the pectoral-fin base. They
can be separated from transitional Labrisomus
by the metamorphic melanophore pattern as well as
the dorsal-fin outline (third-to-last spine less
than a third the length of the longest soft rays
vs. often half or more) and long cirri (about a
pupil width or more vs. short, esp. orbital).
Once they develop a large dark spot at the front
of the spinous dorsal fin, juveniles can be separated
by that feature from most other Malacoctenus
species except M.
gilli, which share the spot but also have
a distinctive ocellus at the rear spinous dorsal
fin, and M.
boehlkei, which have the dorsal-fin spot
ocellated (with yellow) and elevated some distance
above the base of the fin. The inverted-triangle
pattern for which the species is named is not diagnostic
in juveniles- the basic pattern is shared by several
congeners. Juvenile L.
nuchipinnis, L.
conditus, and L.
cricota have a similar dorsal-fin spot and
some may be missing their opercular ocellus or dark
spot; in that case the dorsal-fin outline (with
medium-length posterior spines), a longer third
pelvic-fin ray, and reticulated markings on the
body should distinguish them from M. triangulatus.
Later they diverge markedly in morphology becoming
bulkier with a large mouth and smaller eyes.
dorsal-fin base larval
melanophores
spots at the tip of upper jaw and ethmoid
Barbados HV08, coll.
Henri Valles
Malacoctenus triangulatus
new recruit
16.8 mm SL, DNA-confirmed
ID
St. Thomas, USVI, ST429
Malacoctenus triangulatus
new recruit
14.9 mm SL
persistent larval melanophores
San Blas, Panama, SB83-141
Malacoctenus erdmani
Diagnosis: The modal
fin-ray count of D-XXI,9 A-II,18-19 and P-16 indicates
M. erdmani and fall within the range for
M.
macropus and barely within the range for
M. gilli
(M.
delalandii can match the median-fin ray
count but have fewer than 16 pectoral-fin rays).
(DNA)
Ecology:
The imitator blenny is a tiny labrisomid rarely
noticed by divers. They are not common and prefer
shallow rockier habitats than most of their congeners.
The species is apparently restricted to the Caribbean
Sea and the Bahamas, with no records from Florida,
the Gulf of Mexico, or NE Venezuela down to Brazil
and its offshore islands. Their larvae are uncommon
in collections and, unlike many other reef fishes,
most were collected in the dry season in Panama.
Description: Pre-transitional larvae: Body long,
narrow, and thin with a large round eye, pointed
snout, and relatively small terminal mouth. Long
continuous dorsal and anal fins with a short and
narrow caudal peduncle. Pectoral fins long, reaching
past the vent, and pelvic fins long and thread-like.
On the head there are a pair of large side-by-side
melanophores overlying the midbrain lobes, typically
widely spaced (more than a pupil-width apart), sometimes
with one or two additional small adjacent spots,
but all limited to the the midbrain lobes (nearing
transition, one or a few additional small pseudolarval
spots can develop on the side of the head and anteriorly).
There is a cheek melanophore on each side. There
are no melanophores along the base of the dorsal
or caudal fins. Along the ventral midline there
are notably no melanophores at the isthmus or at
the pelvic-fin base. Along the anal fin there is
a melanophore at the base of each anal-fin soft
ray (and sometimes the second spine), usually sparing
the last ray, followed by a single melanophore placed
just after the last ray (sometimes a second) along
the ventral midline of the caudal peduncle; it is
often slightly more prominent than the preceding
row. Internal melanophores comprise only the basic
complement: the nuchal midline, otic capsule, and
overlying the abdominal organs.
Transitional stage: M. erdmani
larvae in transition develop patches of small surface
melanophores over the head, including a short bar
down from the orbital rim at 6:00 o'clock and an
eye-stripe from the orbital rim to the mid-maxilla.
In addition, the melanophores on the lower rim of
the pectoral-fin base form two patches, the anteriormost
often a vertical line from under the operculum.
Transitional larvae also sometimes develop a few
additional small pseudo-larval melanophores over
the hindbrain lobes. Fine metamorphic melanophores
later extend onto the body. Notably, the cirri that
develop on each side of the head, on the nape, over
the eye, and over the nasal tube are multifid.
Juveniles:M. erdmani juveniles
can be recognized by having a blue-ringed ocellus
forming on the body just below the last few dorsal-fin
spines, often still part of a dark fourth bar on
the body.
Analogues:
Larval M. erdmani (and M.
macropus) can be separated from their congeners
by their light markings, i.e. the absence of melanophores
along the dorsal and caudal-fin bases, none along
the anterior ventral midline forward of the anal
fin, and fewer than 5 melanophores on top of the
head with bare forebrain lobes, i.e. spots only
over the midbrain optic lobes and none over the
forebrain lobes (between the eyes). M.
gilli larvae can appear similar, but have
a deep melanophore at the pelvic-fin insertion (and
typically additional head melanophores and fewer
pectoral-fin rays). M.
aurolineatus larvae can have few melanophores
on the head, but have additional melanophores along
the median-fin bases and at the isthmus and pelvic-fin
insertion. Larval M. erdmani are very similar
to larval M.
macropus in size, shape, and markings. There
are some marking differences, with M. erdmani
larvae always having (at least) a side-by-side pair
of head melanophores while M.
macropus often have a single melanophore.
The arrangement of melanophores on the ventral midline
of the caudal peduncle also differs: M. erdmani
larvae usually have a single, often more prominent,
melanophore placed just after the last fin ray (when
there is a second spot, the two species can overlap),
while M.
macropus usually have two to four evenly-sized
melanophores spaced out along the caudal peduncle;
if one, it is most often placed half-way to the
procurrent caudal-fin rays. Since most M. erdmani
also have fewer anal-fin rays than M.
macropus, the total number of melanophores
in the ventral row is typically two or three fewer
(often 18 vs. 21). Fin-ray counts are generally
different and helpful for separation, however there
is some overlap requiring DNA sequencing for definitive
identification of larvae within the shared range.
The D-XXI,9 A-II,18-19 P-16 combination occurs in
more than half of M. erdmani individuals
but is rare in M.
macropus. Certain counts are indicative
of M. erdmani: 29 total dorsal-fin elements,
i.e. 20 dorsal-fin spines or 21 with only 8 dorsal-fin
rays (below Springer's reported range for M.
macropus) and the frequent combination of
18 anal-fin soft rays and 16 pectoral-fin rays (characteristic
of M. erdmani and rare for M.
macropus). Late larvae also diverge in the
relative length of their dorsal-fin spines: in M.
erdmani the first spine is distinctly longer
than the third-to-last spine, while in M.
macropus the first spine is about the same
length (or less) as the third-to-last spine.
Early transitional M. erdmani larvae are
distinguished by fin-ray counts, persistent larval
melanophores, and their metamorphic melanophore
pattern. During transition M. erdmani diverge
from M.
macropus in the number of cirri that develop:
in M. erdmani the cirri over the eye and
on the nape are bifid or trifid vs. single in M.
macropus. During transition M. erdmani
also diverge from M.
macropus in the relative length of their
dorsal-fin spines; the third-to-last spine is much
shorter than the first spine, while in M.
macropus the several spines before the last
become long, often longer than the first.
Juvenile and adult M. erdmani are best recognized
by the prominent squared-off ocellus ringed in black
and/or blue on the body just below the last few
dorsal-fin spines. Juvenile M.
gilli have an ocellus at the same location,
but it clearly extends onto the fin membranes. Juvenile
M.
macropus have no ocelli and long single
cirri (multiple in M. erdmani).
Ecology:
Rosy blennies are the most common labrisomid in
the Caribbean and are found in large numbers in
all shallow clear-water habitats. They are small,
only one or two inches long, and especially abundant
in the mixed coral, rubble, sand, and seagrass areas
that occupy large areas of hard-bottom shoreline
in the region. The species ranges throughout the
Caribbean, as well as Bermuda, Florida, and the
Gulf of Mexico, but not NE Venezuela or Brazil and
its offshore islands. Their larvae are among the
ten most commonly collected reef fish larvae and
occur in the vast majority of nightlight and plankton-tow
collections.
Description: Pre-transitional larvae: Body long,
narrow, and thin with a large round eye, pointed
snout, and relatively small terminal mouth. Long
continuous dorsal and anal fins with a short and
narrow caudal peduncle. Pectoral fins long, reaching
past the vent, and pelvic fins long and thread-like.
On the head there are either a pair of large side-by-side
melanophores or a single slightly off-center melanophore
overlying the midbrain lobes (about a third of larvae
with the single spot). The paired spots are often
not widely spaced (about a pupil-width apart). Occasionally
there are three or four spots, rarely up to six,
but typically all are clustered over the midbrain
lobes, behind the level of the rear edge of the
orbit (nearing transition, one or a few additional
small pseudolarval spots can develop on the side
of the head and anteriorly). There is a cheek melanophore
on each side. There are no melanophores along the
base of the dorsal or caudal fins. Along the ventral
midline there are notably no melanophores at the
isthmus or at the pelvic-fin base. An occasional
variant (including about half from one large collection)
can have a melanophore along the dorsal midline
of the caudal peduncle or at the base of one or
a few dorsal-fin soft rays and/or on the side of
the body just above the rear anal fin (very rarely
a spot or two at the base of the caudal-fin rays).
Along the anal fin there is a melanophore at the
base of each anal-fin soft ray (and sometimes the
second spine), usually sparing the last ray, followed
by two to four melanophores spaced out along the
ventral midline of the caudal peduncle (occasionally
one or none; if one, it is most often placed half-way
to the procurrent caudal-fin rays). Internal melanophores
comprise only the basic complement: the nuchal midline,
otic capsule, and overlying the abdominal organs.
Transitional stage: M. macropus
larvae in transition develop patches of small surface
melanophores over the head, including a short bar
down from the orbital rim at 5:30 o'clock and an
eye-stripe from the orbital rim to the anterior
third of the maxilla. In addition, the melanophores
on the lower pectoral-fin base form a short line
slanting down and back from under the operculum
or, sometimes, a wider horizontal band across the
lower fin base. Transitional larvae also sometimes
develop a few additional small pseudo-larval melanophores
over the midbrain lobes and transitional juveniles
often show multiple satellite pseudo-larval melanophores
that resemble fragmentation of the large larval
melanophores. Fine metamorphic melanophores later
extend onto the body forming either complex reticulations
in the shape of rows of irregular vertical ovals
or uniform shading, typically denser on the upper
body. Notably, the long cirri that develop on each
side of the head, on the nape, over the eye, and
over the nasal tube are single filaments.
Juveniles:M. macropus juveniles
have highly variable marking patterns: most often
a bicolor pattern of dark above, light below, but
often uniformly-shaded or occasionally even prominently
barred. Their long single cirri (at the nape, over
the eye, and at the nasal tube) on each side are
diagnostic.
Analogues:
Larval M. macropus (and M.
erdmani) can be separated from their congeners
by their light markings, i.e. the absence of melanophores
along the dorsal and caudal-fin bases, none along
the anterior ventral midline forward of the anal
fin, and fewer than 5 melanophores on top of the
head with bare forebrain lobes, i.e. spots only
over the midbrain optic lobes and none over the
forebrain lobes (between the eyes). M.
gilli larvae can appear similar, but have
a deep melanophore at the pelvic-fin insertion (and
typically additional head melanophores and only
20 dorsal-fin spines). M.
aurolineatus larvae can have few melanophores
on the head, but have additional melanophores along
the dorsal and caudal-fin bases and at the isthmus
and pelvic-fin insertion. Larval M.
erdmani can be very similar to larval M.
macropus in size, shape, and markings. Since
M.
erdmani larvae seem to always have a side-by-side
pair of head melanophores, the identification problem
only applies to M. macropus larvae with a
pair of head melanophores, more than half of the
specimens in most samples. The arrangement of melanophores
on the ventral midline of the caudal peduncle differs:
M. macropus usually have two to four melanophores
spaced out along the caudal peduncle after the last
fin ray (if one, it is most often placed half-way
to the procurrent caudal-fin rays), while M.
erdmani larvae usually have a single melanophore
very close to the last fin ray, often slightly more
prominent than the preceding anal-fin row (occasionally
two, then the two species overlap). Since most M.
macropus also have more anal-fin rays than M.
erdmani, the total number of melanophores
in the ventral row is typically two or three more
in M. macropus (often 21 vs. 18). Fin-ray
counts are generally different and helpful for separation,
however there is some overlap requiring DNA sequencing
for definitive identification of larvae within the
shared range. The D-XXI,9 A-II,18-19 P-16 combination
occurs in more than half of M.
erdmani individuals, and is rare in M.
macropus (especially the 18 anal-fin soft rays).
Certain counts are indicative of M.
erdmani: 29 total dorsal-fin elements, i.e.
20 dorsal-fin spines or 21 with only 8 dorsal-fin
rays (rare, and below Springer's reported range,
for M. macropus) and the frequent combination
of 18 anal-fin soft rays and 16 pectoral-fin rays
(characteristic of M.
erdmani and rare for M. macropus).
Late larvae also diverge in the relative length
of their dorsal-fin spines: in M. macropus
the first spine is about the same length (or less)
as the third-to-last spine, while in M.
erdmani the first spine is distinctly longer
than the third-to-last spine.
Early transitional M. macropus larvae are
distinguished by fin-ray counts, persistent larval
melanophores, and their metamorphic melanophore
pattern, i.e. the combination of the 5:30 o'clock
bar, an eye stripe to the front of the maxilla,
and a short line across the pectoral-fin base from
under the operculum (two patches in M.
erdmani). Transitional larvae develop single
cirri on each side of the nape and over the eye,
while congeners, including M.
erdmani, develop multiple cirri when the
new cirri are medium-length. During transition,
M. macropus diverge from their congeners
in the dorsal-fin outline, with the last several
spines long, often longer than the first spine vs.
a particularly short third-to-last spine in the
other species (in this feature, M. macropus
resemble Labrisomus juveniles, although in
all other respects, they are typical Malacoctenus).
Juvenile (and adult) M. macropus have a
variety of marking patterns, from uniformly shaded
(white or rosy) and speckled, to bicolor (dark above,
light below), or even prominently barred, and are
best distinguished by the long single cirri extending
from the nasal tube, eye, and nape vs. multiple,
often bushy, sets of cirri on congeners. When barred,
the bars do not extend over the anal fin and the
two bars under the last dorsal-fin spines meet on
the body to form a prominent V or Y.
Malacoctenus macropus
earlier larva
8.8 mm SL
San Blas, Panama, SB86-045
Malacoctenus macropus
larva
11.6 mm SL
first spine about equal
to third-to-last
San Blas, Panama, SB84-520
Malacoctenus macropus
larvae
11.9 mm transitional
& 11.4 mm SL larva
no anterior ventral midline
melanophores
Belize & San Blas,
Panama, SB86-424
Malacoctenus macropus
larvae
10.8 to 11.7 mm SL
head spot variations
note bare forebrain lobes
San Blas, Panama, SB86-422
Malacoctenus macropus
larva
10.4 mm SL
internal melanophore
pattern
San Blas, Panama, SB86-409
Malacoctenus macropus
transitional larva
12.0 mm SL, DNA-confirmed
ID
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Malacoctenus macropus
transitional larva
11.3 mm SL
single head
melanophore slightly off center
San Blas, Panama, SB86-604
Malacoctenus macropus
transitional larva
11.5 mm SL
additional pseudo-larval
melanophores
San Blas, Panama, SB86-516
Malacoctenus macropus
transitional larva
11.9 mm SL
Glovers Reef, Belize,
coll. Cormac Nolan
Malacoctenus macropus
transitional larva
11.7 mm SL, DNA-confirmed
ID
single nuchal and orbital
cirrus per side
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Malacoctenus macropus
transitional recruit
11.3 mm SL
"fragmented" larval head
melanophores
San Blas, Panama, SB81-069
Malacoctenus
macropus
transitional recruits
11.8, 11.4, and 11.5
mm SL
shaded or patterned;
note long single
cirri and long last few dorsal-fin spines
San Blas, Panama, SB92
Malacoctenus macropus
new recruit
14.5 mm SL, DNA-confirmed
ID
uncommon barred variant
St. Thomas, USVI, ST429
Malacoctenus boehlkei
Diagnosis: Fin-ray
counts of D-XX,13 or XXI,11-12 or XXII,11 (a mode
of 33 dorsal-fin elements) and modal A-II,22 with
15 pectoral-fin rays indicates Malacoctenus boehlkei.
Few congeners match the high dorsal-fin ray count;
only a rare M.
triangulatus shares the D-XX,13 and a very
rare M.
macropus has as many as D-XXIII,10 and/or
A-II,22. Labrisomus filamentosus share the
high median-fin ray count, but have only 13 pectoral-fin
rays (and a quite different morphology). L.
haitiensis barely overlaps the count, with
a rare D-XXII,11 and/or A-II,22. (DNA)
Ecology:
The diamond blenny is a tiny blenny typically found
on coral walls and slopes below 30 feet; it is the
only deeper-water Malacoctenus species and
often shelters among anemone tentacles. The species
ranges from Florida, the S. Gulf of Mexico, and
the Bahamas across the Caribbean Sea, but not NE
Venezuela or Brazil and its offshore islands. Their
larvae are unknown or unrecognized in collections.
Description: (larvae
unknown, description inferred from transitional
juvenile) Larvae: Body long, narrow, and thin
with a large round eye, pointed snout, and relatively
small terminal mouth. Long continuous dorsal and
anal fins with a short and narrow caudal peduncle.
Pectoral fins long, reaching past the vent, and
pelvic fins very long and thread-like. On the head
there are several large and small melanophores per
side, over both the fore- and midbrain lobes. Along
the anal fin there is a melanophore at the base
of each anal-fin soft ray.
Juveniles:M. boehlkei recruits
develop a distinctive yellow-ringed ocellus on the
first three dorsal-fin-spine membranes, not contacting
the body. Their two longest pelvic-fin rays are
greatly extended, reaching to the mid-anal fin.
Analogues:
Once they develop the characteristic ocellated spot
at the front of the spinous dorsal fin, recruits
can be separated from all other labrisomid species.
M.
triangulatus and M.
gilli both have a dark non-ocellated spot
on the first dorsal-fin spines, located lower on
the fin and extending onto the body.
Malacoctenus boehlkei
trans juvenile
15.8 mm SL, DNA-confirmed
ID
St. Thomas, USVI, ST951
Labrisomus
presented in order of increasing dorsal-fin elements
Labrisomus albigenys
Diagnosis: The modal
fin-ray count of D-XVIII,11 A-II,18 and P-13 indicates
Labrisomus albigenys and L.
nigricinctus. This fin-ray count falls within
the lower range for Malacoctenus
aurolineatus and M.
versicolor, as well as for many of the remaining
18 and 19-spined Labrisomus. (DNA)
Ecology:
The white-cheek blenny is a rare species, with few
collections and no live photographs. They are found
sporadically throughout the Caribbean Sea, as well
as in the Bahamas and the Yucatan, but not farther
into the Gulf of Mexico or in Florida, Bermuda,
NE Venezuela, or Brazil and its offshore islands.
Their larvae are rare in collections.
Description: Pre-transitional larvae: Body long,
moderately narrow, and thin with a large round eye,
pointed snout, and medium-sized terminal mouth.
Long continuous dorsal and anal fins with a very
short and relatively narrow caudal peduncle. Pectoral
fins long, reaching past the vent, and pelvic fins
long and thread-like. On the head there are a pair
of large side-by-side melanophores overlying the
midbrain lobes and smaller melanophores, either
one off-center or a pair, over the forebrain lobes.
The cheek melanophores are apparently absent on
the three larvae identified. There are no melanophores
along the base of the dorsal or caudal fins, although
one specimen has one and two melanophores on the
sides of the body just lateral to the base of the
last dorsal-fin spines (perhaps transitional). Along
the ventral midline there is a melanophore at the
isthmus and deep at the pelvic-fin base. Along the
anal fin there is a melanophore at the base of each
anal-fin soft ray (sometimes at the second spine
as well). In addition, there is a variably-placed
large melanophore near the base of the 14th anal-fin
soft ray, sometimes at the base of the ray and extending
into the musculature, but sometimes located subsurface
in the body some distance away from the base of
the fin ray. There are no melanophores along the
ventral midline of the caudal peduncle. Internal
melanophores comprise only the basic complement:
the nuchal midline, otic capsule, and overlying
the abdominal organs.
Transitional stage: L. albigenys
larvae in transition develop diffuse areas of
fine surface melanophores over the top and side
of the head, not in defined patches. A pale bar
develops across the cheek from 4-5 o'clock on the
orbital rim. The species apparently develops metamorphic
melanophores over the head first, unlike most Labrisomus
species. Cirri develop on each side of the head,
on the nape (relatively stubby), over the eye (delayed),
and over the nasal tube. One larva caught at a light
at night had a fully-scaled body before developing
metamorphic melanophores.
Juveniles: L. albigenys juveniles
have no opercular ocellus (or dark spot) and usually
an anterior dorsal-fin spot. They have a prominent
pale patch on the cheek below the eye (4-6 o'clock
from the orbit) outlined by two dark lines.
Analogues:
Larval L. albigenys can be distinguished
from congeners by the prominent enlarged melanophore
in or near the posterior anal-fin base series (internal
into the musculature or sub-surface on the body
near the series). A similar kind of melanophore
can be found in the anal-fin base series of some
Starksia
larvae, however they all have fewer than 9 dorsal-fin
soft rays. L. albigenys larvae share their
small size, relatively small terminal mouth, and
dorsal-fin outline (short first spine and short
posterior spines), with the larvae of L.
nigricinctus and some Malacoctenus.
L. albigenys larvae are very similar to larval
L.
nigricinctus in both morphology and fin-ray
counts, and can best be separated by the enlarged
or deep anal-fin melanophore and perhaps by having
no melanophores on the ventral caudal peduncle (vs.
one). L. albigenys larvae are separated from
Malacoctenus by melanophore pattern and fin-ray
counts. Only M.
gilli larvae occasionally share the pattern
of a pair over the midbrain lobes and one or a pair
over the forebrain lobes, but M.
gilli larvae have no isthmus melanophores.
M.
erdmani and M.
macropus can have similarly few head spots,
but they are over the midbrain lobes only and they
have no anterior ventral midline spots. Only rare
individuals of M.
aurolineatus and M.
versicolor share the low median and pectoral-fin
ray counts of L. albigenys and both have
different arrangements of head melanophores: the
former with a midline melanophore over the midbrain
lobes and additional melanophores along the soft
dorsal and caudal-fin bases and the latter with
many more head spots. Larvae of the remaining Labrisomus
species have more head spots, at least five
in a U- or V-shape, relatively longer posterior
dorsal-fin spines (vs. soft rays), and are generally
larger at transition.
TransitionalL. albigenys larvae
are distinguished by fin-ray counts, persistent
larval melanophores, and their metamorphic melanophore
pattern, i.e. the metamorphic melanophores on the
head are diffusely distributed with only a pale
bar across the cheek. Transitional L.
nigricinctus are unknown, but probably do
not share the persistent enlarged or deep anal-fin
melanophore and the head melanophore distribution.
L. albigenys are separated from transitional
Malacoctenus by having stubs of nuchal cirri
and not having clearly delineated patches of metamorphic
melanophores on the head. The remaining Labrisomus
species develop metamorphic melanophores over the
head and body simultaneously, have more larval head
melanophores, and relatively longer posterior dorsal-fin
spines.
L. albigenys juveniles have a uniform dark
head with a pale patch on the cheek under the eye,
usually outlined by two dark lines, although a similar
pattern is common to lesser degrees among many Labrisomus
species. Juvenile Malacoctenus all have well-outlined
and distinctive head markings. The dorsal-fin outline,
with the short first spine and very short penultimate
spine relative to the soft rays, separates L.
albigenys juveniles from several other Labrisomus
species (except L.
nigricinctus, L.
guppyi and, to a degree, L.
gobio). The XVIII,11 or fewer dorsal-fin
elements also helps to distinguish less-marked L.
albigenys juveniles from occasional lightly-marked
juveniles of other Labrisomus species, although
a few can range down to overlap. The absence of
a dark spot or ocellus on the operculum separates
L. albigenys from the other 18-spined species:
L.
nigricinctus and L.
nuchipinnis, L.
conditus, and L.
cricota (which all also have longer snouts,
more than two-thirds the bony orbit diameter). The
19- and 20-spined Labrisomus species share
the blunter snout with juvenile L. albigenys
(less than two-thirds the bony orbit diameter),
but only rarely have so few median fin-rays. L.
albigenys early juveniles also have been described
to have only two nasal cirri and one long simple
orbital cirrus per side (more or branched in the
other Labrisomus and Malacoctenus
species, except M.
macropus).
Labrisomus albigenys
larva
12.7 mm SL
very short first dorsal
spine
internal & surface melanophore deep to rear
anal fin, no cheek spot
San Blas, Panama, SB86-1008
Labrisomus albigenys
larva
12.1 mm SL
surface melanophore deep
to rear anal fin
San Blas, Panama, SB81-105
Labrisomus albigenys
transitional larva
12.2 mm SL, DNA-confirmed
ID
first dorsal spine short,
lower than third spine and much less than long
soft-rays
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Labrisomus albigenys
holotype
16.0 mm SL, from Beebe
& Tee-Van 1928
note only 10 soft dorsal-fin
rays
Port-au-Prince, Haiti,
9 May 1927
Labrisomus nigricinctus
Diagnosis: The modal
fin-ray count of D-XVIII,11 A-II,18 and P-13 indicates
Labrisomus nigricinctus and L.
albigenys. This fin-ray count falls within
the lower range for Malacoctenus
aurolineatus and M.
versicolor, as well as for many of the remaining
18 and 19-spined Labrisomus species. (DNA)
Note: This larval identification is pending DNA
confirmation, but is likely from the process of
elimination: the larva differs in several ways from
the L.
albigenys series described above (the L.
albigenys identification is based on the
fin-ray counts and a very different DNA sequence
from adult L. nigricinctus from the same
site). The remaining species of Labrisomus
and Malacoctenus
that could possibly share the low fin-ray count
of this larva have different larvae, all clearly
identified here with DNA-sequence confirmation.
The remarkably small size of the larva (for Labrisomus)
matches the observation by Springer (1959) that
juvenile L. nigricinctus as small as 12 mm
SL are collected on the reef.
Ecology:
The spotcheek blenny is a small blenny found mostly
in tidepools and shallow eroded reef and thus rarely
noticed underwater. They are found throughout the
Caribbean, as well as Florida, the Gulf of Mexico,
and the Bahamas, but not NE Venezuela or Brazil
and its offshore islands. The species is common
only in clear water and high-energy sites: the outer
Keys in Florida, the Bahamas and Antilles, and the
barrier reef and atolls of Belize. Their larvae
are rare in collections.
Description: Pre-transitional larvae: Body long,
moderately narrow, and thin with a medium round
eye, pointed snout, and relatively small terminal
mouth. Long continuous dorsal and anal fins with
a short and narrow caudal peduncle. Pectoral fins
long, reaching past the vent, and pelvic fins long
and thread-like. On the head there are two small
off-center melanophores overlying the midbrain lobes.
There is a cheek melanophore on each side. There
are no melanophores along the dorsal or caudal-fin
bases. Along the ventral midline there is a melanophore
at the isthmus and deep at the pelvic-fin base.
Along the anal fin there is a melanophore at the
base of each anal-fin soft ray except the last,
which is closely followed by a single larger melanophore
along the ventral midline of the caudal peduncle.
Internal melanophores comprise only the basic complement:
the nuchal midline, otic capsule, and overlying
the abdominal organs.
Juveniles: L. nigricinctus
juveniles are quite distinctive with a prominent
round opercular ocellus, a pointed snout with a
small terminal mouth, no spot on the first few dorsal-fin
spines, and sometimes narrow dark bars on the body
that extend onto the dorsal fin.
Analogues: L. nigricinctus larvae share fin-ray counts
and some morphology with L.
albigenys, but differ by having a melanophore
along the ventral midline of the caudal peduncle,
one or two cheek melanophores, and no deep melanophore
associated with the anal-fin row. These two species
are intermediate in size and appearance between
Labrisomus and Malacoctenus,
but, notably, have lower fin-ray counts than all
Malacoctenus
except the rare individual of M.
versicolor or M.
aurolineatus. Larvae of M.
versicolor can be distinguished by their
relatively long first dorsal-fin spines (vs. short
in the two Labrisomus species) and more numerous
head spots. M.
aurolineatus larvae also have a longer first
dorsal-fin spine and have a set of additional larval
melanophores (along the soft dorsal-fin and caudal-fin
bases and internally over the vertebral column).
M.
macropus, M.
erdmani and M.
gilli larvae have similar light markings,
but no melanophore at the isthmus (or the pelvic-fin
base in the former two) and higher fin-ray counts.
The remaining Malacoctenus species have many
more head spots. The remaining Labrisomus
larvae are larger with larger mouths and usually
with many more melanophores over the cranium, dorsal
and caudal-fin bases, and internally over the vertebral
column.
Juvenile L. nigricinctus are separated from
other Labrisomus species by the small mouth,
pointed snout, and well-delineated round opercular
ocellus. In morphology they appear more similar
to Malacoctenus
juveniles, however no Malacoctenus
share the obvious ocellus on the operculum. Juvenile
L.
nuchipinnis, L.
guppyi, and L.
filamentosus can share the opercular ocellus,
but are distinguished by less-pointed snouts, larger
mouths, and indistinct bars. The 19- and 20-spined
Labrisomus species all have a much blunter
snout, less than two-thirds the bony orbit diameter.
Ecology:
The true hairy blenny is the largest member of
its family in the region (adults can reach more
than six inches long) and they are the most commonly
encountered species of Labrisomus in the
Caribbean Sea (but not Florida). They occupy a
variety of shallow-water habitats, but are usually
found in mixed coral and rocky substrates. The
recent discovery of two additional allied cryptic
species described from Brazil but widespread in
the region complicates identifications within
the group. L. nuchipinnis is widely distributed:
from Bermuda and the Bahamas, as well as the Caribbean
Sea to mainland Brazil. They are replaced on Noronha
off Brazil by L.
conditus and share the Brazilian coast
with L.
cricota. DNA sequencing results thus far
show that outside of Brazil and Florida, L.
nuchipinnis well outnumber the cryptic species
(or are the only species). L. nuchipinnis
barcode DNA sequences vary little over the entire
region from Bermuda to Brazil. In Florida, the
vast majority of hairy blennies sequenced prove
to be L.
conditus, with a few L.
cricota. Of course, it is likely that
all three species could be found in S. Florida
where Caribbean and Florida lineages of many reef
fishes often overlap.
True hairy blennies vary greatly in markings
and colors, with some overlap in appearance with
the cryptic species, although many smaller specimens
(and all juveniles?) show the sharp and narrow
white rim around a rounded opercular ocellus that
likely confirms the ID as L. nuchipinnis.
However, when the ocellus is not round, or not
well-delineated by a thin white ring, or the white
ring is broken, or there is no ocellus, the marking
does not separate adults of the three species
(I have found DNA-confirmed adult L. nuchipinnis
can have any or all of these patterns, including
a mix of any version on each side of the fish).
Their larvae are occasional in collections.
Description: Pre-transitional larvae: Body long,
narrow, and thin, with a medium eye, pointed snout,
and terminal medium-sized mouth. Long continuous
dorsal and anal fins with a short and narrow caudal
peduncle. Pectoral fins long, reaching past the
vent, and pelvic fins long and thread-like; the
third pelvic-fin ray is about 3/4 the length of
the second. There are some small spines along the
rim of the preoperculum that no longer project as
larvae approach and complete transition. The full
complement of large melanophores on the top of the
head typically consists of a row of three on each
side of the head, usually in a narrow-U, i.e. the
spots get closer to the dorsal midline anteriorly;
quite often there are just five (rarely 7) in a
V with a single melanophore at the vertex at or
near the midline of the forebrain (note that in
both cases the rear side-by-side pair are typically
more widely spaced than the middle pair). Often
there is an additional near-surface melanophore
at the midline behind the mid-brain lobes, completing
a narrow-O (not the deep nuchal midline melanophore).
There can sometimes be one or, uncommonly, a few
smaller additional melanophores. There are no melanophores
just behind the tip of the upper jaw, but there
can be a small melanophore at the anterior nostril
on each side. There is a cheek melanophore on each
side. There is a prominent melanophore, or sometimes
a few, on the inner aspect of the cleithrum visible
within the gill cavity on each side. Melanophores
run along the base of all of the soft dorsal-fin
rays and some of the dorsal-fin spines, typically
including some anterior spines, usually starting
at spine 8 (can be at 5 or even 2), then 11-12,
and from 14 rearward. A few individuals develop
a small melanophore along the dorsal midline of
the caudal peduncle. A small melanophore is often
located on the body at the lateral midline on the
caudal peduncle. A vertical line of melanophores
develops along the base of the caudal-fin segmented
rays (first proximal, then distal) and thin linear
melanophores are spaced out along each side outlining
the full-length of the three lower caudal-fin segmented
rays (occasionally a few along the upper three rays
as well). Along the ventral midline there are melanophores
at the isthmus and deep behind the pelvic-fin base.
There is occasionally a small melanophore or two
at the surface on the ventral aspect of the abdomen,
but no extensive speckling of the peritoneum is
visible from below. Along the anal fin there is
a melanophore at the base of each anal-fin soft
ray, followed by one, sometimes two, along the ventral
midline of the caudal peduncle. A row of internal
melanophores overlies the vertebral column, spaced
about every third vertebra, along the mid- and rear
body, continuing onto the caudal peduncle. Additional
internal melanophores include those at the nuchal
midline, otic capsule, and overlying the abdominal
organs.
Transitional stage: L. nuchipinnis
larvae develop metamorphic melanophores over the
head and body simultaneously. The head is mostly
uniformly speckled or reticulated with fine melanophores
while the body is uniformly reticulated with a network
of thin lines not separated into bars. As in other
Labrisomus, the prominent head melanophores
begin to shrink, spread, or fragment into smaller
spots, or narrow into short lines (usually the middle
pair); later they are obscured by underlying speckling
and overlying shading.
Juveniles: L. nuchipinnis
juveniles develop a pattern of reticulations and
dark bars on the body, an opercular ocellus, usually
fully rounded with a complete thin white rim, and
a dark spot on the first three dorsal-fin spine
membranes. Juvenile blennies of this group can vary
greatly in the degree of these dark markings; some
light individuals intensify their white spots and
fade, or sort of pixelate, their melanocytes, while
others are darkly-reticulated and some can be almost
uniformly dark.
Analogues:
Pretransitional L. nuchipinnis are probably
identical to the larvae of the cryptic species L.
conditus and L.
cricota. Although the first dorsal-fin spine
is shorter than the third in juvenile and adult L. nuchipinnis
and L.
conditus vs. longer in L.
cricota, DNA-confirmed L. nuchipinnis
larvae can have equal-length anterior dorsal-fin
spines and it is likely that the fin-spine differences
only emerge at or after transition. Larvae of the
other 18-spined species, L.
albigenys and L.
nigricinctus, have many fewer melanophores
on the head and usually none at the dorsal and caudal-fin
bases. L.
haitiensis larvae share the slim morphology
with L. nuchipinnis larvae as well as the
row of melanophores along the spinous dorsal-fin
base including some of the first 10 spines, and,
to varying degrees, most of the other markings of
larval L. nuchipinnis, but they can be distinguished
by a short and inconspicuous third pelvic-fin ray,
a pair of melanophores behind the tip of the upper
jaw (absent on L. nuchipinnis larvae), extensive
speckling of the ventral abdominal viscera visible
through the abdominal wall, the absence of inner
cleithral melanophores, and higher fin-ray counts.
The remaining Labrisomus species usually
have an unmarked spinous dorsal-fin base or, at
most, melanophores extending only to some of the
spines of the rear half of the spinous dorsal-fin.
Those with more melanophores typically have the
pair of melanophores behind the tip of the upper
jaw (absent on L. nuchipinnis larvae) as
well as additional melanophores on top of the head,
multiple in each quadrant (vs. the basic U or V-pattern
on L. nuchipinnis), and rarely have the inner
cleithral melanophores. Lightly marked specimens
can be problematic because they are often missing
the pair at the tip of the jaw and share the basic
U or V-pattern of L. nuchipinnis; in that
case they can be distinguished by the absence of
the inner cleithral melanophores and the absence
of the melanophores lining the lower caudal-fin
segmented rays (or, rarely, a very few at the proximal
ends of the rays).
L. nuchipinnis larvae can resemble large
Malacoctenus larvae since they are relatively
slim and have smaller mouths than some other large
Labrisomus species. Nevertheless, the row
of melanophores along some of the spinous dorsal-fin
base separates L. nuchipinnis from all Malacoctenus
larvae other than the occasional variant specimen
of M.
triangulatus. The latter, however, do not
have melanophores outlining the caudal-fin segmented
rays or the central caudal peduncle spot and usually
have more numerous and graded-size head spots (as
well as higher median-fin ray counts, a short third
pelvic-fin ray, and a different dorsal-fin outline,
with short posterior spines).
Transitional L. nuchipinnis larvae are distinguished
by fin-ray counts, persistent larval melanophores,
and their metamorphic melanophore pattern, i.e.
a uniform pattern of fine reticulations. The other
Labrisomus species develop uniform shading
or bars, except for the two allied cryptic species
which share the reticulations of L. nuchipinnis:
transitional L.
cricota may be distinguished by the first
dorsal-fin spine being longer than the third, and
transitional L.
conditus probably cannot be separated until
the oval dark spot develops on the operculum. L.
nuchipinnis are also separated from transitional
Malacoctenus larvae by having shorter cirri
(especially orbital), the fine metamorphic melanophores
on the head mostly diffusely distributed (vs. in
discrete patches) and developing at the same time
as body markings (vs. head first), and the dorsal-fin
outline.
Juvenile L. nuchipinnis are separated from
most other labrisomids by the long snout, anterior
dorsal fin spot, and a thin-edged rounded opercular
ocellus. Juveniles of L.
conditus and L.
cricota share the anterior dorsal-fin spot
and body reticulations and bars, but apparently
do not have a round well-delineated opercular ocellus:
juvenile L.
conditus have an oval and not sharply-outlined
ocellus on the operculum and L.
cricota have a diffusely broad and orange
edge to the opercular ocellus and the first two
dorsal-fin spines distinctly longer than the third
(vs. equal or shorter in the other two species).
Two other Labrisomus have a well-outlined
ocellus, L.
nigricinctus and L.
filamentosus, but the former have narrow
bars and a very pointed snout and the latter have
the first three dorsal-fin spines greatly extended
and higher median-fin ray counts. Two other species
have a less well-outlined ocellus: L.
guppyi, with no dorsal-fin spot, a blunt
snout, and 19 dorsal-fin spines; and L.
haitiensis, with the opercular spot not
outlined as an ocellus, a blunt snout, an inconspicuous
third pelvic-fin ray, and 20 dorsal-fin spines.
The 19- and 20-spined Labrisomus species
all have a blunt snout, less than two-thirds the
bony orbit diameter. Juvenile Malacoctenus
all have pointier snouts with smaller mouths, have
very short posterior dorsal-fin spines (except M.
macropus), and none have a well-outlined
opercular ocellus.
Labrisomus nuchipinnis
larva
19.2 mm SL
first spines short and
equal length
San Blas, Panama, SB81-200
Labrisomus nuchipinnis
larvae
U, V, and O patterned
head spots
Panama and Barbados
Labrisomus nuchipinnis
larva
18.9 mm SL, DNA-confirmed
ID
full complement of tail
melanophores
San Blas, Panama, SB84-624
Labrisomus nuchipinnis
larva
17.5 mm SL
inner cleithral spots,
long third pelvic ray
San Blas, Panama, SB81-200
Labrisomus nuchipinnis
larva
18.3 mm SL, DNA-confirmed
ID
first spine slightly
shorter than third
San Blas, Panama, SB81-242
Labrisomus nuchipinnis
transitional larva
19.3 mm SL, DNA-confirmed
ID
reticulations, no ocellus
yet
first spine about equal
to third
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Labrisomus nuchipinnis
transitional juv.
20.3 mm SL, DNA-confirmed
ID
round opercular ocellus,
anterior dorsal fin spot, equal first 3 spines,
long snout
Ecology:
The masquerader hairy blenny was recently described
as the endemic hairy blenny from the Brazilian
offshore island of Fernando de Noronha by Sazima,
Carvalho-Filho, Gasparini & Sazima (2009).
However, barcode DNA surveys show that the same
species has been collected in Panama and Yucatan
and, indeed, most specimens sequenced in Florida
are L. conditus as well. Although the original
description notes some marking differences from
the two other cryptic species and shorter nuchal
cirri in L. conditus, it is uncertain whether
these apply throughout the wide shared range (or
to juveniles). L. conditus are supposed
to have a less well-developed opercular ocellus
with an incomplete and wide orange rim and a profusion
of small blue spots over the head and body. However,
true hairy blennies (with DNA sequence confirmation)
vary greatly in markings and colors and some adults
can overlap in appearance with the cryptic species
(although many have a narrowly-delineated rounded
opercular ocellus). A transitional juvenile Panamanian
L. conditus has a distinctive oval-shaped
opercular ocellus that is not sharply outlined,
however the consistency of this mark is uncertain.
L. conditus larvae are unknown or unrecognized
in collections.
Description:
(based on the transitional specimen, larval patterns
are identical to L.
nuchipinnis)
Body long, narrow, and thin, with a medium eye,
pointed snout, and terminal medium-sized mouth.
Long continuous dorsal and anal fins with a short
and narrow caudal peduncle. Pectoral fins long,
reaching past the vent, and pelvic fins long and
thread-like; the third pelvic-fin ray is about 3/4
the length of the second. There are some small spines
along the rim of the preoperculum that no longer
project as larvae approach and complete transition.
The full complement of large melanophores on the
top of the head typically consists of a row of three
on each side of the head, usually in a narrow-U,
i.e. the spots get closer to the dorsal midline
anteriorly; quite often there are just five (rarely
7) in a V with a single melanophore at the vertex
at or near the midline of the forebrain (note that
in both cases the rear side-by-side pair are typically
more widely spaced than the middle pair). Often
there is an additional near-surface melanophore
at the midline behind the mid-brain lobes, completing
a narrow-O (not the deep nuchal midline melanophore).
There can sometimes be one or, uncommonly, a few
smaller additional melanophores. There are no melanophores
just behind the tip of the upper jaw, but there
can be a small melanophore at the anterior nostril
on each side. There is a cheek melanophore on each
side. There is a prominent melanophore, or sometimes
a few, on the inner aspect of the cleithrum visible
within the gill cavity on each side. Melanophores
run along the base of all of the soft dorsal-fin
rays and some of the dorsal-fin spines, typically
including some anterior spines, usually starting
at spine 8 (can be at 5 or even 2), then 11-12,
and from 14 rearward. A few individuals develop
a small melanophore along the dorsal midline of
the caudal peduncle. A small melanophore is often
located on the body at the lateral midline on the
caudal peduncle. A vertical line of melanophores
develops along the base of the caudal-fin segmented
rays (first proximal, then distal) and thin linear
melanophores are spaced out along each side outlining
the full-length of the three lower caudal-fin segmented
rays (occasionally a few along the upper three rays
as well). Along the ventral midline there are melanophores
at the isthmus and deep behind the pelvic-fin base.
There is occasionally a small melanophore or two
at the surface on the ventral aspect of the abdomen,
but no extensive speckling of the peritoneum is
visible from below. Along the anal fin there is
a melanophore at the base of each anal-fin soft
ray, followed by one, sometimes two, along the ventral
midline of the caudal peduncle. A row of internal
melanophores overlies the vertebral column, spaced
about every third vertebra, along the mid- and rear
body, continuing onto the caudal peduncle. Additional
internal melanophores include those at the nuchal
midline, otic capsule, and overlying the abdominal
organs.
Transitional stage: L. conditus
larvae develop metamorphic melanophores over the
head and body simultaneously. The head is mostly
uniformly speckled or reticulated with fine melanophores
while the body is uniformly reticulated with a network
of thin lines not separated into bars. As in other
Labrisomus, the prominent head melanophores
begin to shrink, spread, or fragment into smaller
spots, or narrow into short lines (usually the middle
pair); later they are obscured by underlying speckling
and overlying shading.
Juveniles:L. conditus develop
a pattern of reticulations and dark bars on the
body, a dark oval ocellus without a sharp outline
on the operculum, and a dark spot on the first three
dorsal-fin spine membranes. Juvenile blennies of
this group can vary greatly in the degree of these
dark markings; some light individuals intensify
their white spots and fade, or sort of pixelate,
their melanocytes, while others are darkly-reticulated
and some can be almost uniformly dark.
Analogues:
Pretransitional L. conditus are probably
identical to the larvae of the cryptic species L.
nuchipinnis and L.
cricota. Although the first dorsal-fin spine
is shorter than the third in adult L. conditus
and L.
nuchipinnis vs. longer in L.
cricota, it is likely that the fin spine
differences only emerge at or after transition.
L. conditus larvae can resemble large Malacoctenus
larvae since they have thinner heads (side-to-side),
smaller eyes, and smaller mouths than most other
large Labrisomus species. Nevertheless, the
melanophore row along the base of some of the spinous
dorsal fin separates L. conditus larvae from
all Malacoctenus larvae other than the occasional
variant specimen of M.
triangulatus. The latter, however, do not
have melanophores outlining the caudal-fin segmented
rays or the central caudal peduncle spot and usually
have more numerous and graded-size head spots (as
well as higher median-fin ray counts, a short third
pelvic-fin ray, and a different dorsal-fin outline,
with short posterior spines). L.
haitiensis larvae have a similar slim morphology
and share the row of melanophores along some of
the spinous dorsal-fin base and, to varying degrees,
most of the other markings of larval L. conditus,
but they can be distinguished by a short and inconspicuous
third pelvic-fin ray, a pair of melanophores behind
the tip of the upper jaw (apparently absent on L.
conditus larvae), and higher fin-ray counts.
Transitional L. conditus larvae are distinguished
by fin-ray counts, persistent larval melanophores,
and their metamorphic melanophore pattern, i.e.
a uniform pattern of fine reticulations. They are
also separated from transitional Malacoctenus
larvae by having shorter cirri (especially orbital),
the fine metamorphic melanophores on the head mostly
diffusely distributed (vs. in discrete patches),
as well as developing at the same time as body markings
(vs. head first), and the dorsal-fin outline (relatively
longer posterior spines). The other Labrisomus
species develop uniform shading or bars, except
for the two allied cryptic species which share the
reticulations of L. conditus: transitional
L.
cricota may be distinguished by the first
dorsal-fin spine being longer than the third, and
transitional L.
nuchipinnis probably can only be separated
when they develop their well-outlined rounded opercular
ocellus.
Juvenile L. conditus are separated from
most other labrisomids by the long snout, anterior
dorsal fin spot, dark reticulations on the body,
and an oval opercular ocellus that is not sharply
outlined. L.
nuchipinnis, L.
nigricinctus, and L.
filamentosus have a well-outlined rounded
ocellus. Other Labrisomus juveniles with
a dark spot on the operculum include the cryptic
species L.
cricota, which share most markings but have
a diffusely broad and orange edge to the opercular
ocellus and the first two dorsal-fin spines distinctly
longer than the third (vs. equal or shorter in the
other two species);
L. guppyi, with no dorsal-fin spot; and
L.
haitiensis, with no dorsal-fin spot and
a short third pelvic fin ray. The 19- and 20-spined
Labrisomus species can be separated by higher
fin-ray counts and blunter snouts, less than two-thirds
the bony orbit diameter.
Labrisomus conditus
transitional juvenile
22.5 mm SL, DNA-confirmed
ID
oval opercular ocellus,
anterior dorsal fin spot, equal length first
3 spines
Ecology:
The mock hairy blenny was recently described as
endemic to the mainland coast of Brazil by Sazima,
Gasparini & Leao de Moura (2002). However,
my barcode DNA surveys show that the same species
has been collected in Panama, Dominica, and Bahamas,
and there is a match to a specimen collected in
Florida as well. The first two dorsal-fin spines
are longer than the third in L. cricota,
and, although the original description notes some
marking differences and longer nuchal cirri in
L. cricota, it is uncertain whether these
latter characters are consistent throughout the
wide range. L. cricota adults have a less
well-developed opercular ocellus with a wide and
diffuse orange rim, however true hairy blennies
(with DNA sequence confirmation) vary greatly
in markings and colors and some adults can overlap
in appearance with the cryptic species (although
many have a narrowly-delineated rounded opercular
ocellus). L. cricota larvae are rare in
collections.
Description:
(based on the transitional specimen, larval patterns
are identical to L.
nuchipinnis)
Body long, narrow, and thin, with a medium eye,
pointed snout, and terminal medium-sized mouth.
Long continuous dorsal and anal fins with a short
and narrow caudal peduncle. Pectoral fins long,
reaching past the vent, and pelvic fins long and
thread-like; the third pelvic-fin ray is about 3/4
the length of the second. There are some small spines
along the rim of the preoperculum that no longer
project as larvae approach and complete transition.
The full complement of large melanophores on the
top of the head typically consists of a row of three
on each side of the head, usually in a narrow-U,
i.e. the spots get closer to the dorsal midline
anteriorly; quite often there are just five (rarely
7) in a V with a single melanophore at the vertex
at or near the midline of the forebrain (note that
in both cases the rear side-by-side pair are typically
more widely spaced than the middle pair). Often
there is an additional near-surface melanophore
at the midline behind the mid-brain lobes, completing
a narrow-O (not the deep nuchal midline melanophore).
There can sometimes be one or, uncommonly, a few
smaller additional melanophores. There are no melanophores
just behind the tip of the upper jaw, but there
can be a small melanophore at the anterior nostril
on each side. There is a cheek melanophore on each
side. There is a prominent melanophore, or sometimes
a few, on the inner aspect of the cleithrum visible
within the gill cavity on each side. Melanophores
run along the base of all of the soft dorsal-fin
rays and some of the dorsal-fin spines, typically
including some anterior spines, usually starting
at spine 8 (can be at 5 or even 2), then 11-12,
and from 14 rearward. A few individuals develop
a small melanophore along the dorsal midline of
the caudal peduncle. A small melanophore is often
located on the body at the lateral midline on the
caudal peduncle. A vertical line of melanophores
develops along the base of the caudal-fin segmented
rays (first proximal, then distal) and thin linear
melanophores are spaced out along each side outlining
the full-length of the three lower caudal-fin segmented
rays (occasionally a few along the upper three rays
as well). Along the ventral midline there are melanophores
at the isthmus and deep behind the pelvic-fin base.
There is occasionally a small melanophore or two
at the surface on the ventral aspect of the abdomen,
but no extensive speckling of the peritoneum is
visible from below. Along the anal fin there is
a melanophore at the base of each anal-fin soft
ray, followed by one, sometimes two, along the ventral
midline of the caudal peduncle. A row of internal
melanophores overlies the vertebral column, spaced
about every third vertebra, along the mid- and rear
body, continuing onto the caudal peduncle. Additional
internal melanophores include those at the nuchal
midline, otic capsule, and overlying the abdominal
organs.
Transitional stage: L. cricota
larvae develop metamorphic melanophores over the
head and body simultaneously. The head is mostly
uniformly speckled or reticulated with fine melanophores
while the body is uniformly reticulated with a network
of thin lines not separated into bars. As in other
Labrisomus, the prominent head melanophores
begin to shrink, spread, or fragment into smaller
spots, or narrow into short lines (usually the middle
pair); later they are obscured by underlying speckling
and overlying shading.
Juveniles:L. cricota develop
a pattern of reticulations and dark bars on the
body, a dark spot on the first three dorsal-fin
spine membranes, and a diffusely broad orange-outlined
opercular ocellus. Juvenile blennies of this group
can vary greatly in the degree of these dark markings;
some light individuals intensify their white spots
and fade, or sort of pixelate, their melanocytes,
while others are darkly-reticulated and some can
be almost uniformly dark.
Analogues:
Pretransitional L. cricota are probably identical
to the larvae of the cryptic species L.
nuchipinnis and L.
conditus. As larvae approach transition,
the first dorsal-fin spine becomes longer than the
third in L. cricota vs. shorter or equal
in L.
nuchipinnis and L.
conditus. L. cricota larvae can resemble
large Malacoctenus larvae since they have
thinner heads (side-to-side), smaller eyes, and
smaller mouths than most other large Labrisomus
species. Nevertheless, the melanophore row along
the base of some of the spinous dorsal fin separates
L. cricota larvae from all Malacoctenus
larvae other than the occasional variant specimen
of M.
triangulatus. The latter, however, do not
have melanophores outlining the caudal-fin segmented
rays or the central caudal peduncle spot and usually
have more numerous and graded-size head spots (as
well as higher median-fin ray counts, a short third
pelvic-fin ray, and a different dorsal-fin outline,
with short posterior spines). L.
haitiensis larvae have a similar slim morphology
and share the row of melanophores along some of
the spinous dorsal-fin base and, to varying degrees,
most of the other markings of larval L. cricota,
but they can be distinguished by a short and inconspicuous
third pelvic-fin ray, a pair of melanophores behind
the tip of the upper jaw (apparently absent on L.
cricota larvae), and higher fin-ray counts.
Transitional L. cricota larvae are distinguished
by fin-ray counts, persistent larval melanophores,
and their metamorphic melanophore pattern, i.e.
a uniform pattern of fine reticulations. They are
also separated from transitional Malacoctenus
larvae by having shorter cirri (especially orbital),
the fine metamorphic melanophores on the head mostly
diffusely distributed (vs. in discrete patches)
and developing at the same time as body markings
(vs. head first), and the dorsal-fin outline. The
other Labrisomus species develop uniform
shading or bars, except for the two allied cryptic
species which share the reticulations of L. cricota:
transitional L.
nuchipinnis and L.
conditus are best distinguished by the first
dorsal-fin spine being shorter than or equal to
the third.
Juvenile L. cricota are separated from most
other labrisomids by the anterior dorsal fin spot
and a diffusely broad orange-outlined opercular
ocellus. L.
nuchipinnis, L.
nigricinctus, and L.
filamentosus have a well-outlined rounded
ocellus. Other Labrisomus juveniles with
a dark spot on the operculum include the cryptic
species L.
conditus, which have most of the same markings
but the first dorsal-fin spine is shorter than or
equal to the third (vs. longer); L.
guppyi, with no dorsal-fin spot; and L.
haitiensis, with no dorsal-fin spot and
a short third pelvic fin ray. The 19- and 20-spined
Labrisomus species can be separated by higher
fin-ray counts and blunter snouts, less than two-thirds
the bony orbit diameter.
Labrisomus cricota
transitional larva
17.0 mm SL, DNA-confirmed
ID
first dorsal spine slightly
longer than third
faded melanophores not illustrated
San Blas, Panama, SB84-526a
Labrisomus cricota
transitional juvenile
22.6 mm SL, DNA-confirmed
ID
specimen with white marks
predominant
first dorsal-fin spine
longer than third
San Blas, Panama, SB83-150
Labrisomus gobio
Diagnosis: The modal
fin-ray count of D-XIX,11 A-II,19 and P-13 is shared
by the 19-spined trio of L. gobio, L.
guppyi, and L.
kalisherae, as well as by Malacoctenus
aurolineatus (but the latter usually with
14 pectoral-fin rays). Note that almost 90% of L.
gobio individuals have 19 dorsal-fin spines.
The fin-ray count falls within the upper or lower
range for most other Labrisomus species and
both M.
delalandii and M.
gilli. (DNA)
Ecology:
The palehead blenny is a somewhat common small blenny
found generally in shallow mixed coral habitats.
The species is widespread in the region, found in
Florida, the S. Gulf of Mexico, the Bahamas, and
across the Caribbean Sea except NE Venezuela. Unfortunately,
the 19, 20, and 21-spined Labrisomus are
very similar morphologically and are frequently
confused in guidebooks and photographs. All six
species share the blunt-headed and goggle-eyed appearance,
at least as juveniles, i.e. the snout length (bony
orbit to jaw tip) is less than two-thirds of the
orbit diameter. Adult palehead blennies are recognized
in the field mostly by the absence of an opercular
ocellus, the dark bars not extending onto the fins,
and relatively even-length dorsal-fin spines. Additional
characters often cited are an "unmarked"
lower head and the lower portion of the bars on
the body being lighter and narrower than the upper
parts, although the variation in the intensity of
markings in the group make these unreliable. Juveniles
are particularly difficult to separate, since the
distinguishing markings for each species are either
not well-developed or shared by other species at
this early stage (but see description below). L.
gobio larvae are rare in collections.
Description:
(pre-transitional larvae have not been identified,
but are likely identical among the 19-spined Labrisomus,
see L.
guppyi)
Transitional stage: L. gobio
larvae in transition develop metamorphic melanophores
over the head and body simultaneously. The head
is mostly uniformly speckled with fine melanophores;
on the body there is a series of about 9 dark bars,
alternating lighter and darker, with unpigmented
bands between them.
Juveniles:L. gobio juveniles
have prominent dark bars on the body that do not
extend onto the fins, with the second body bar (under
the 10-15th dorsal-fin spines) widening on the upper
body to touch the bases of about 5 or 6 spines.
The last dark bar, on the tail, ends as a straight
vertical line or slight crescent. Although there
can be a dark spot on the operculum, it is not an
ocellus with a thin rim. The first dorsal-fin spine
is relatively long, about half the body depth at
the dorsal-fin origin, and similar in length to
the mid-fin spines.
Analogues:
The juveniles of the 19- and 20-spined Labrisomus
are blunt-headed, compared to the 18-spined Labrisomus
(other than L.
albigenys), i.e. the snout length (bony
orbital margin to the tip of the closed jaw) is
less than two-thirds the bony orbit diameter. Small
juveniles of the blunt-headed Labrisomus
are difficult to identify to species, especially
since some species described with opercular ocelli
can have only a diffuse dark spot on the operculum
when young. In addition, those species described
without ocelli can often have an outlined dark spot
on the operculum (but not a thin-rimmed ocellus)
as juveniles.
Juvenile L. gobio are best identified by
the dark bars on the body that do not extend onto
the fins and the second body bar widening to reach
about 5 or 6 dorsal spine bases (vs. 3 or 4 in other
species). They can be further separated from L.
guppyi juveniles who have not yet developed
a distinct opercular ocellus by a longer first dorsal-fin
spine (2 to 2.5 into body depth at the dorsal-fin
origin vs. 2.5 to 3) and fewer dark bands on the
pelvic fins (often faint and no more than 4). Juvenile
L.
kalisherae share the long first dorsal-fin
spine but have the pigment on the base of the caudal
fin outlining three light spots vs. ending as a
mostly straight vertical line in L. gobio
(and L.
guppyi). Juvenile L.
bucciferus and L.
haitiensis also have three outlined spots
on the caudal-fin base (and even longer first dorsal-fin
spines and higher fin-ray counts). L.
albigenys are best distinguished by their
very short first dorsal-fin spine and lower fin-ray
counts. The remaining Labrisomus juveniles
are distinguished from L. gobio by longer
snouts, opercular ocelli, and fin-ray counts, i.e.
L.
nigricinctus and L.
nuchipinnis (as well as L.
conditus and L.
cricota, both with less distinct ocelli),
and L.
filamentosus (blunt-snouted but with longer
first dorsal-fin spines).
Labrisomus gobio
juvenile
25.3 mm SL, DNA-confirmed
ID
bars not on fins, widening
second bar
vertical line at caudal-fin base
first dorsal spine 2.5 into body depth
St. Thomas, USVI ST955
Labrisomus guppyi
Diagnosis: The modal
fin-ray count of D-XIX,11 A-II,19 and P-13 is shared
by the 19-spined trio of L. guppyi, L.
gobio, and L.
kalisherae, as well as by Malacoctenus
aurolineatus (but usually with 14 pectoral-fin
rays). Note that almost 90% of L. guppyi
individuals have 19 dorsal-fin spines. The fin-ray
count falls within the upper or lower range for
most other Labrisomus species and both M.
delalandii and M.
gilli. (DNA)
Ecology:
The mimic blenny is a somewhat common small blenny
found mostly under rocks and in shallow complex
limestone and mixed coral habitats. The species
is widespread in the region, found in Florida, the
S. Gulf of Mexico, the Bahamas, and across the Caribbean
Sea except NE Venezuela. Unfortunately, the 19,
20, and 21-spined Labrisomus are very similar
morphologically and are frequently confused in guidebooks
and photographs. All six species share the blunt-headed
and goggle-eyed appearance, at least as juveniles,
i.e. the snout length (bony orbit to jaw tip) is
less than two-thirds of the orbit diameter. Adult
mimic blennies are recognized in the field mostly
by a distinct opercular ocellus, well-outlined and
typically with an orange posterior rim, and a relatively
short first dorsal-fin spine. Juveniles are particularly
difficult to separate, since the distinguishing
markings for each species are either not well-developed
or shared by other species at this early stage (but
see description below). L. guppyi larvae
are uncommon in collections.
Description: Pre-transitional larvae: Body long,
narrow, and thin with a medium eye, pointed snout,
and terminal medium-sized mouth. Larvae can have
the normal body shape or develop a hunched-over
appearance (tip of snout below the lateral midline
of the body). Long continuous dorsal and anal fins
with a short and narrow caudal peduncle. Pectoral
fins long, reaching past the vent, and pelvic fins
long and thread-like; the third pelvic-fin ray is
about 3/4 the length of the second. There are some
small spines along the rim of the preoperculum that
no longer project as larvae approach transition.
The complement of melanophores on the top of the
head is usually a narrow-U, i.e. a row of three
on each side of the head with the spots closer to
the dorsal midline anteriorly, plus a near-surface
melanophore at the midline behind the mid-brain
lobes (not the deep nuchal midline melanophore),
completing a narrow-O (note that the rear side-by-side
pair are typically more widely spaced than the middle
pair). Usually there are additional equally large
melanophores over some or all four quadrants, i.e.
each lobe of the fore- and mid-brain. On more marked
larvae, there is a prominent melanophore on each
side just behind the tip of the upper jaw (this
pair is often absent on lightly marked larvae).
There is a cheek melanophore on each side. Melanophores
typically run along the base of all of the soft
dorsal-fin rays, and, on more heavily marked larvae,
along some of the posterior dorsal-fin spines as
well; however some lightly-marked larvae have only
a few soft-dorsal fin base spots or even rarely
none. A vertical line of melanophores develops along
the base of the caudal-fin segmented rays (first
proximal, then distal), sometimes few and mostly
on the upper half, or rarely none, on lightly marked
larvae. Some larvae can have one or two linear melanophore
streaks outlining the mid-length of one or two lower
caudal-fin segmented rays (most have none). Along
the ventral midline there are melanophores at the
isthmus and deep behind the pelvic-fin base. There
is no extensive speckling of the peritoneum visible
through the ventral abdominal wall, but some more
heavily marked larvae can show some peritoneal speckling
on the upper sidewalls of the abdomen. Along the
anal fin there is a melanophore at the base of each
anal-fin soft ray, followed by none to three along
the ventral midline of the caudal peduncle. A row
of internal melanophores overlies the vertebral
column, spaced about every third vertebra, along
the mid- and rear body, continuing onto the caudal
peduncle. Additional internal melanophores include
those at the nuchal midline, otic capsule, and overlying
the abdominal organs.
Transitional stage: L. guppyi
larvae in transition develop metamorphic melanophores
over the head and body simultaneously, a mostly
uniformly speckling of fine melanophores, although
some transitional larvae develop an indistinct pattern
of dark bars.The full complement of larval melanophores,
especially multiple spots over each quadrant of
the forebrain and midbrain as well as the pair behind
the tip of the upper jaw are present at transition.
Juveniles:L. guppyi juveniles
have dark bars on the body that extend onto the
dorsal-fin membranes, with the second body bar (under
the 10-15th dorsal-fin spines) covering the bases
of about 4 spines. The last dark bar on the tail
ends as a straight vertical line or a slight crescent.
Early juveniles may have only a diffuse dark spot
on the operculum, but they rapidly develop an ocellus
with a thin orange rim on the posterior edge. The
first dorsal-fin spine is notably short, only a
third or less of the body depth at the dorsal-fin
origin and well shorter than the mid-fin spines.
Analogues:
The larvae of the 19-spined Labrisomus are
likely identical, although, as they approach transition,
L. guppyi larvae should develop their shorter
first dorsal-fin spine.
Lightly-marked larvae of L. guppyi missing
the pair of melanophores behind the tip of the upper
jaw can be distinguished from larvae of L.
nuchipinnis, L.
conditus, and L.
cricota, by the absence of melanophores
along the base of the anterior half of the spinous
dorsal fin, especially the first ten spines, and
the absence of melanophores outlining the lower
segmented rays of the caudal fin (also many of the
latter three species have only a single melanophore
over the forebrain quadrants (V of 5) vs. more on
L. guppyi). L.
albigenys and L.
nigricinctus larvae are very lightly-marked,
missing most of the melanophore complement of L.
guppyi. Among the Malacoctenus larvae,
only M.
aurolineatus and a rare M.
triangulatus share the melanophores along
the base of the soft dorsal fin; the former have
a distinctive head melanophore pattern and the latter
are distinguished by a short third pelvic-fin ray,
many head melanophores varying from tiny to large,
and a dorsal-fin outline with a short third-to-last
spine. Other Malacoctenus larvae may resemble
the rare L. guppyi without dorsal midline
melanophores, but they all have fewer head melanophores
not in the U or O pattern and/or dorsal-fin outlines
with short posterior spines and often different
fin-ray counts.
L. guppyi larvae with the pair of melanophores
behind the tip of the upper jaw can be distinguished
from L.
bucciferus larvae by a shorter first dorsal-fin
spine (as they approach transition) and usually
one fewer dorsal-fin spine (possibly also by additional
small head melanophores, if present, over all quadrants
vs. only forebrain). L.
haitiensis larvae differ in having abdominal
speckling, melanophores at the base of dorsal-fin
spines 8 or 9, longer pelvic fins with a short third
ray, and additional dorsal-fin spines.
Transitional L. guppyi larvae are distinguished
by fin-ray counts, persistent larval melanophores,
and their uniform speckling of metamorphic melanophores.
The pair of melanophores behind the tip of the upper
jaw is shared only by the other 19- and 20+-spined
species, L.
gobio, L.
kalisherae, L.
bucciferus, and L.
haitiensis, all of which have a different
dorsal-fin outline, with longer first spines (about
two to 2.5 times in the body depth at the dorsal-fin
origin vs. 3 times or more, and about the same length
as the the mid-fin spines vs. well shorter). Transitional
L.
haitiensis develop a prominent pattern of
ovals and bars. The transitional larvae of L.
nuchipinnis, L.
conditus, and L.
cricota develop reticulated lines over the
body (vs. uniform speckling). Transitional L.
guppyi larvae are separated from transitional
Malacoctenus larvae by the uniform markings
over the head and body (developing at the same time),
short cirri, and the dorsal-fin outline.
The juveniles of the 19- and 20-spined Labrisomus
are blunt-headed, compared to the 18-spined Labrisomus
(other than L.
albigenys), i.e. the snout length (bony
orbital margin to the tip of the closed jaw) is
less than two-thirds the bony orbit diameter. Small
juveniles of the blunt-headed Labrisomus
are difficult to identify to species, especially
since some species described with opercular ocelli
can have only a diffuse dark spot on the operculum
when young. In addition, those species described
without ocelli can often have an outlined dark spot
on the operculum (but not a thin-rimmed ocellus)
as juveniles.
Juvenile L. guppyi who have not yet developed
a distinct opercular ocellus are difficult to separate
from L.
gobio juveniles: the key differences are
a shorter first dorsal-fin spine (about a third
the body depth at the dorsal-fin origin and well
shorter than the mid-fin spines vs. about half and
slightly shorter, about 90%), the dark bars on the
body extend onto the fins in L. guppyi (vs.
not onto the fins) and the mid-body bar covers about
3 or 4 spine bases (vs. widening to reach 5 or 6).
Juvenile L.
kalisherae, as well as L.
bucciferus and L.
haitiensis (both with higher fin-ray counts),
are separated from L. guppyi by having distinctly
longer first dorsal-fin spines (about two into the
body depth and the same length or a little shorter
than the mid-fin spines vs. well shorter) and the
pigment on the caudal peduncle ending by outlining
three light spots vs. a mostly straight vertical
line in both L. guppyi and L.
gobio. Juvenile L.
albigenys share the dorsal-fin outline of
L. guppyi (relatively short first and penultimate
dorsal-fin spines) and are best distinguished by
the absence of an opercular spot or ocellus and
their lower fin-ray counts. After juvenile L.
guppyi develop an opercular ocellus with a thin
rim (at least on the posterior edge), they can be
distinguished from other ocellated species: from
L.
nigricinctus and L.
nuchipinnis (as well as L.
conditus and L.
cricota, both with less distinct ocelli)
by the blunt snout and from L.
haitiensis (which can have a similar
opercular spot) and L.
filamentosus by the short first dorsal-fin
spine (and from all by fin-ray counts).
Labrisomus guppyi
larvae
14.7 mm SL
San Blas, Panama SB80-102
Labrisomus guppyi
larvae
16.0 and 14.1 mm SL
hunched body and straight
San Blas, Panama SB81-105
Labrisomus guppyi
larvae
12.5-14.7 mm SL
range of head melanophore
patterns
San Blas, Panama
Labrisomus guppyi
? transitional larva
17.7 mm SL, cannot r/o
gobio
spots nr ascending process
of premaxilla
diffusely speckled head and body
multiple spots over all head quadrants
Barbados HV08, coll.
Henri Valles
Labrisomus guppyi
? transitional larva
16.1 mm SL, cannot r/o
gobio
short first dorsal spine,
well shorter than mid-spines, diffusely speckled
head and body, multiple spots over all cranium
quadrants, pair at jaw tip
San Blas, Panama, SB81-050
Labrisomus guppyi
transitional larva
17.0 mm SL, DNA-confirmed
ID
v. short first dorsal
spine, 3.5 in body depth & well shorter than
mid-spines
diffusely speckled head and body
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Labrisomus guppyi
juvenile
22.6 mm SL, DNA-confirmed
ID
obscure opercular ocellus
bars onto fins, second body bar not wide
vertical line at caudal-fin base
first dorsal spine three into body depth
St. Thomas, USVI ST9430
Labrisomus kalisherae
Diagnosis: The modal
fin-ray count of D-XIX,11 A-II,19 and P-13 is shared
by the 19-spined trio of L. kalisherae, L.
gobio, and L.
guppyi, as well as by Malacoctenus
aurolineatus (but usually with 14 pectoral-fin
rays). Note that almost 90% of L. kalisherae
individuals have 19 dorsal-fin spines. The fin-ray
count falls within the upper or lower range for
most other Labrisomus species and both M.
delalandii and M.
gilli. (DNA)
Ecology:
The downy blenny is an uncommon small blenny found
mostly in inshore shallow habitats, particularly
associated with algae and rubble. The species ranges
from Florida and the S. Gulf of Mexico along the
continental coast and across the Southern Caribbean
to NE Venezuela (where it is the only 19- or 20-spined
Labrisomus species), Noronha, and mainland
Brazil. However, it is common only in cooler and/or
turbid-water areas such as the inshore reefs of
Belize and Florida, Venezuela, and Brazil. It appears
to be replaced by L.
bucciferus in warmer clear-water areas such
as the Bahamas, the Antilles, and the offshore barrier
reef and atolls in Belize. Unfortunately, the 19,
20, and 21-spined Labrisomus are very similar
morphologically and are frequently confused in guidebooks
and photographs. All six species share the blunt-headed
and goggle-eyed appearance, at least as juveniles,
i.e. the snout length (bony orbit to jaw tip) is
less than two-thirds of the orbit diameter. Adult
downy blennies are recognized in the field mostly
by the absence of an opercular ocellus, intensely
spotted fins, especially the anal fin, relatively
long first two dorsal-fin spines (and, on close
inspection, only two submandibular pores). Juveniles
are particularly difficult to separate, since the
distinguishing markings for each species are either
not well-developed or shared by other species at
this early stage (but see description below). L.
kalisherae larvae are unknown or unrecognized
in collections.
Description:
(pre-transitional larvae have not been identified,
but are likely identical among the 19-spined Labrisomus,
see L.
guppyi)
Transitional stage: Transitional
L. kalisherae larvae have not been identified.
Juveniles:L. kalisherae
juveniles have dark bars on the body that extend
onto the fins. The last dark bar ends on the caudal-fin
base outlining three light spots. Although there
can be a dark spot on the operculum, it is not an
ocellus with a thin rim. The ventral and pectoral
fins have dark spotting, with 5 or 6 distinct dark
bands along the pelvic fin-rays. The first dorsal-fin
spine is relatively long, about half the body depth
at the dorsal-fin origin, and similar in length
or slightly shorter than the mid-fin spines.
Analogues:
Transitional L. kalisherae larvae likely
differ from the other 19-spined Labrisomus
only by the length of the first dorsal-fin spine,
which should be relatively long, not distinctly
shorter than the fourth to sixth spines.
The juveniles of the 19- and 20-spined Labrisomus
are blunt-headed, compared to the 18-spined Labrisomus
(other than L.
albigenys), i.e. the snout length (bony
orbital margin to the tip of the closed jaw) is
less than two-thirds the bony orbit diameter. Small
juveniles of the blunt-headed Labrisomus
are difficult to identify to species, especially
since some species described with opercular ocelli
can have only a diffuse dark spot on the operculum
when young. In addition, those species described
without ocelli can often have an outlined dark spot
on the operculum (but not a thin-rimmed ocellus)
as juveniles.
L. kalisherae are almost identical to L.
bucciferus by appearance, although fortunately
the latter typically have 20 dorsal-fin spines.
The adult character of the number of pores at the
submandibular symphysis does not apply to juveniles.
The marking differences are quite subtle, with L.
kalisherae having more spotted ventral and pectoral
fins, especially more dark bands along the pelvic
fin-rays (5 to 7 vs. 4) and L.
bucciferus are more likely to show short
dark lines radiating from the rear orbital rim and
numerous small pale spots over the operculum. Juvenile
L. kalisherae are best separated from L.
gobio and L.
guppyi by having the pigment on the caudal-fin
base outlining three light spots (vs. ending as
a mostly straight vertical line) and by having distinctly
longer first two dorsal-fin spines (vs. L.
guppyi, i.e. about half the body depth at
the dorsal-fin origin and longer than or equal to
the 4th), often with membranes opacified with white
or colors (vs. both species). In addition, L.
gobio differs in having the dark bars on
the body not extending onto the fins and L.
guppyi rapidly develops an opercular ocellus.
Juvenile L.
haitiensis (and L.
bucciferus) share the relatively long first
dorsal-fin spines (often with white-shaded membranes)
with L. kalisherae, but L.
haitiensis have longer pelvic fins (reaching
past the anal-fin origin) with an inconspicuous
short third pelvic-fin ray, often a dark opercular
spot, and higher fin-ray counts. L.
albigenys are best distinguished by their
short first dorsal-fin spine and lower fin-ray counts.
The remaining Labrisomus juveniles are distinguished
by their opercular ocelli, longer snouts, and fin-ray
counts i.e. L.
nigricinctus and L.
nuchipinnis (as well as L.
conditus and L.
cricota, both with less distinct ocelli),
and L.
filamentosus (blunt-snouted but with longer
first dorsal-fin spines).
Labrisomus kalisherae
juvenile
47.8 mm SL, DNA-confirmed
ID
bars onto fins, three
pale spots at tail
first dorsal spine twice into body depth &
similar length to mid-fin spines
5 or 6 dark bands on pelvic-fin rays
spotted pectoral-fin rays
Fernando de Noronha,
Brazil, FN01
Labrisomus bucciferus
Diagnosis: The modal
fin-ray count of D-XX,11 A-II,20 and P-13 indicates
Labrisomus bucciferus, as well as some chaenopsids
of Emblemariopsis.
Note that almost 90% of L. bucciferus individuals
have 20 dorsal-fin spines. This fin-ray count falls
within the upper range for L.
gobio and L.
kalisherae and the lower range for L.
haitiensis (usually with 14 pectoral-fin
rays), and overlaps the range for Malacoctenus
delalandii, M.
gilli, M
aurolineatus, and M.
triangulatus. (DNA)
Ecology:
The puffcheek blenny is a common small blenny found
mostly in shallow rocky, seagrass, and mixed coral
habitats. The species is widespread in the region,
found in Florida, the Dry Tortugas in the Gulf of
Mexico, Bermuda, the Bahamas, and across the Caribbean
Sea except NE Venezuela. However, they are common
only in warmer and clear-water areas such as the
Bahamas, the Antilles, and the offshore barrier
reef and atolls in Belize and notably uncommon in
Florida and absent from most of the Gulf of Mexico
and Venezuela. It appears to be replaced by L.
kalisherae in cooler and/or turbid-water
areas such as Florida and Venezuela and the inshore
reefs of Belize. Unfortunately, the 19, 20, and
21-spined Labrisomus are very similar morphologically
and are frequently confused in guidebooks and photographs.
All six species share the blunt-headed and goggle-eyed
appearance, at least as juveniles, i.e. the snout
length (bony orbit to jaw tip) is less than two-thirds
of the orbit diameter. Adult puffcheek blennies
are recognized in the field mostly by the lack of
an opercular spot or ocellus, dark lines behind
the eye and pale spotting over the operculum, and
relatively long first two dorsal-fin spines. Juveniles
are particularly difficult to separate, since the
distinguishing markings for each species are either
not well-developed or shared by other species at
this early stage (but see description below). L.
bucciferus larvae are occasional in collections.
Description:
(pre-transitional larvae have not been identified,
but the description is based on the transitional
larvae; lightly marked larvae may be missing some
of the melanophore complement) Pre-transitional larvae: Body long,
narrow, and thin with a medium eye, pointed snout,
and terminal medium-sized mouth. Larvae can have
the normal body shape or develop a hunched-over
appearance (tip of snout below the lateral midline
of the body). Long continuous dorsal and anal fins
with a short and narrow caudal peduncle. Pectoral
fins long, reaching past the vent, and pelvic fins
long and thread-like; the third pelvic-fin ray is
about 3/4 the length of the second. There are some
small spines along the rim of the preoperculum that
no longer project as larvae approach transition.
The full complement of large melanophores on the
top of the head typically consists of a row of three
on each side of the head, usually in a narrow-U,
i.e. the spots get closer to the dorsal midline
anteriorly, usually with one to a few additional
spots over the forebrain lobes (note that the rear
side-by-side pair are often more widely spaced than
the middle pair). There is a prominent melanophore
on each side just behind the tip of the upper jaw.
There is a cheek melanophore on each side. Melanophores
run along the base of all of the soft dorsal-fin
rays as well as some of the posterior dorsal-fin
spines. A vertical line of melanophores develops
along the base of the caudal-fin segmented rays
curving around to the base of the larger procurrent
rays. Along the ventral midline there are melanophores
at the isthmus and deep behind the pelvic-fin base.
There is no extensive speckling of the peritoneum
visible through the ventral abdominal wall, but
some more heavily marked larvae can show some peritoneal
speckling on the upper sidewalls of the abdomen.
Along the anal fin there is a melanophore at the
base of each anal-fin soft ray, followed by none
to three along the ventral midline of the caudal
peduncle. A row of internal melanophores overlies
the vertebral column, spaced about every third vertebra,
along the mid- and rear body, continuing onto the
caudal peduncle. Additional internal melanophores
include those at the nuchal midline, otic capsule,
and overlying the abdominal organs.
Transitional stage: L. bucciferus
larvae in transition develop metamorphic melanophores
over the head and body simultaneously. The head
and body are mostly uniformly speckled with fine
melanophores, although some transitional larvae
develop an indistinct pattern of dark bars.
Juveniles:L. bucciferus
juveniles have dark bars on the body that extend
onto the fins. The last dark bar ends on the caudal-fin
base outlining three light spots. There is usually
no dark spot or ocellus on the operculum, which
is typically covered with small pale spots. The
dark lines radiating from the rear orbit are usually
prominent. The anal and pectoral fins have a few
dark bands, but are not heavily spotted, and the
pelvic fin-rays usually have 3-4 dark bands. The
first two dorsal-fin spines become long, more than
half the body depth at the dorsal-fin origin, and
equal or longer then the mid-fin spines.
Analogues:
Based on the transitional stage, pretransitional
L. bucciferus larvae share the pair of melanophores
behind the tip of the upper jaw with the 19-spined
species and L.
haitiensis. Unlike those species, the extra
melanophores in addition to the basic U or O head
spot pattern may be limited to the forebrain lobes
vs. over all quadrants (it remains to be confirmed
if this pattern is consistent). Otherwise, they
share melanophore patterns with the 19-spined
species and the (usually) extra dorsal-fin spine
may then be the only distinction from L.
gobio and L.
kalisherae (L.
guppyi should have a shorter first dorsal-fin
spine). L.
haitiensis larvae usually differ in having
abdominal speckling, melanophores at the base of
dorsal-fin spines 8 or 9, and longer pelvic fins
with a short third ray.
The larvae of the chaenopsid blennies of Emblemariopsis
can be distinguished by having a gap between the
3rd and fourth dorsal-fin spines, only 3 procurrent
caudal-fin rays (vs. 6 or more), a slimmer body
shape, and many fewer melanophores.
Transitional L. bucciferus larvae are distinguished
by fin-ray counts, persistent larval melanophores,
and their uniform speckling of metamorphic melanophores.
The pair of melanophores behind the tip of the upper
jaw is shared only by the 19-spined species (L.
gobio, L.
guppyi, and L.
kalisherae) and L.
haitiensis. The transitional larvae of L.
gobio and L.
kalisherae have not been identified, but
likely can only be distinguished by the fin-ray
count. Transitional L.
haitiensis as well as L.
nuchipinnis, L.
conditus, and L.
cricota develop reticulated patterns over
the body (vs. uniform speckling). The dorsal-fin
outline separates transitional L. bucciferus
from many congeners: the first spine is relatively
long, about two to 2.5 times in the body depth at
the dorsal-fin origin, and about the same length
as the the mid-fin spines (vs. distinctly shorter,
3 times or more into body depth, in L.
nuchipinnis, L.
conditus, L.
albigenys, L.
nigricinctus, and
L. guppyi,
and well shorter than the mid-fin spines in the
latter three species). Transitional L. bucciferus
larvae are separated from transitional Malacoctenus
larvae by the uniform markings over the head and
body (developing at the same time), short cirri,
and the dorsal-fin outline.
The juveniles of the 19- and 20-spined Labrisomus
are blunt-headed, compared to the 18-spined Labrisomus
(other than L.
albigenys), i.e. the snout length (bony
orbital margin to the tip of the closed jaw) is
less than two-thirds the bony orbit diameter. Small
juveniles of the blunt-headed Labrisomus
are difficult to identify to species, especially
since some species described with opercular ocelli
can have only a diffuse dark spot on the operculum
when young. In addition, those species described
without ocelli can often have an outlined dark spot
on the operculum (but not a thin-rimmed ocellus)
as juveniles.
Juvenile L. bucciferus are difficult to
separate from L.
kalisherae by appearance, although fortunately
L.
kalisherae typically have only 19 dorsal-fin
spines. The adult character of the number of pores
at the submandibular symphysis does not apply to
juveniles. The marking differences are quite subtle,
with L. bucciferus having less spotted ventral
and pectoral fins and fewer dark bands along the
pelvic fin-rays (3 or 4 vs. 5 or 6), and more likely
to show short dark lines radiating from the rear
orbital rim and numerous small pale spots over the
operculum. Juvenile L.
haitiensis (and L.
kalisherae) share the relatively long first
dorsal-fin spines (often with white-shaded membranes)
with L. bucciferus, but L.
haitiensis have distinctly longer pelvic
fins (reaching past the anal-fin origin and with
6-7 bands) with a short and inconspicuous third
pelvic-fin ray, and often a dark opercular spot.
Juvenile
L. bucciferus are best separated from L.
gobio and L.
guppyi by having the pigment on the caudal-fin
base outlining three light spots (vs. ending as
a mostly straight vertical line) and higher fin-ray
counts. In addition, L.
gobio differ in having the dark bars on
the body not extending onto the fins and L.
guppyi have a much shorter first dorsal-fin
spine (less than a third of the body depth at the
dorsal-fin origin vs. half or more) and rapidly
develop an opercular ocellus. L.
albigenys are best distinguished by their
short first dorsal-fin spine and lower fin-ray counts.
The remaining Labrisomus juveniles are distinguished
by their opercular ocelli, longer snouts, and fin-ray
counts i.e. L.
nigricinctus and L.
nuchipinnis (as well as L.
conditus and L.
cricota, both with less distinct ocelli),
and L.
filamentosus (blunt-snouted but with longer
first dorsal-fin spines).
Labrisomus bucciferus
transitional larva
17.0 mm SL, DNA-confirmed
ID
hunched body, uniform
speckling
first dorsal spine 2.5 into body depth &
similar length to mid-fin spines
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Labrisomus bucciferus
transitional juvenile
15.9 mm SL, DNA-confirmed
ID
hunched body, bars onto
fins
first dorsal spine twice into body depth &
similar length to mid-fin spines
fragmented larval melanophores
St. Thomas, USVI, ST9-430
Labrisomus bucciferus
juvenile
22.0 mm SL, DNA-confirmed
ID
bars onto fins, three
pale spots at tail
first dorsal spine twice into body depth &
similar length to mid-fin spines
3-4 dark bands on short pelvic-fin rays
Ecology:
The reef blenny is a somewhat common small blenny
found mostly on well-developed coral reefs at moderate
depths. The species is found in Florida, the Bahamas,
the S. Gulf of Mexico, and across the Caribbean
Sea except NE Venezuela. Unfortunately, the 19,
20, and 21-spined Labrisomus are very similar
morphologically and are frequently confused in guidebooks
and photographs. All six species share the blunt-headed
and goggle-eyed appearance, at least as juveniles,
i.e. the snout length (bony orbit to jaw tip) is
less than two-thirds of the orbit diameter. Adult
reef blennies are recognized in the field mostly
by a longer pelvic fin, reaching the anal-fin origin
(when straightened) and with an inconspicuous short
third ray, as well as relatively long first two
dorsal-fin spines (and, on close inspection, 21
dorsal-fin spines and 14 pectoral-fin rays). Although
they are often described as having an unmarked operculum,
many have a prominent opercular spot, sometimes
even outlined as an ocellus. Juveniles can be difficult
to identify, since the distinguishing markings for
each species are either not well-developed or shared
by other species at this early stage (but see description
below). L. haitiensis larvae are occasional
in collections.
Description: Pre-transitional larvae: Body long,
narrow, and thin with a large eye, pointed snout,
and terminal medium-sized mouth. Long continuous
dorsal and anal fins with a short and narrow caudal
peduncle. Pectoral fins long, reaching past the
vent, and pelvic fins long and thread-like, reaching
more than two-thirds the way to the anal-fin origin;
notably the third pelvic-fin ray is well less than
half the length of the second. The typical complement
of large melanophores on the top of the head consists
of a row of three on each side of the head, usually
in two crescents forming a wide-U, i.e. the middle
side-by-side pair are often more widely spaced than
the rear pair. Sometimes there is an additional
near-surface melanophore at the midline behind the
mid-brain lobes, completing a wide-O (not the deep
nuchal midline melanophore). Some larvae develop
one or a few additional smaller melanophores and
an uncommon "peppered head" variant has
numerous additional small melanophores over the
mid-brain lobes. All larvae have a pair of melanophores
behind the tip of the upper jaw, located adjacent
to the base of the ascending process of the premaxilla
on each side. Some larvae develop additional small
melanophores around the pair. There is a cheek melanophore
on each side. Melanophores run along the base of
all of the soft dorsal-fin rays and some of the
dorsal-fin spines, usually including spines 8 or
9 and 10, then 13-15, and then 17 rearward, but
often all spines after the 8th, and occasionally
only starting at the last two spines. A vertical
line of melanophores develops along the base of
the caudal-fin segmented rays curving around to
the base of the larger procurrent rays. Rare individuals
have thin linear melanophores outlining the length
of the upper three and lower three caudal-fin segmented
rays. Along the ventral midline there are melanophores
at the isthmus and deep behind the pelvic-fin base.
There is usually extensive speckling of the peritoneum
visible both through the lateral abdominal wall
as well as along the ventral abdominal aspect. Along
the anal fin there is a melanophore at the base
of each anal-fin soft ray, followed by one, sometimes
none or two, along the ventral midline of the caudal
peduncle. A row of internal melanophores overlies
the vertebral column along the mid- and rear body,
spaced about every third vertebra, usually continuing
onto the caudal peduncle. Additional internal melanophores
include those at the nuchal midline, otic capsule,
and overlying the abdominal organs.
Transitional stage: L. haitiensis
larvae develop metamorphic melanophores in a distinctive
pattern of isolated loops and rings with vertical
extensions. As transition progresses most of the
larval melanophores on top of the head disappear
and often only a few fragments are apparent.
Juveniles:L. haitiensis
juveniles have dark bars on the body that extend
onto the fins. The last dark bar ends on the caudal-fin
base outlining three light spots. There can be a
dark spot, often with white edging (but not an obvious
complete ocellus), on the operculum. L. haitiensis
have long pelvic fins, usually reaching past the
anal-fin origin, with a distinctively short and
inconspicuous third pelvic-fin ray (well less than
half the length of the second).
Analogues:
There is some overlap between the melanophore patterns
of larval L. haitiensis and the
19-spined species and L.
bucciferus; however, L. haitiensis
larvae apparently always have the combination of
the pair behind the tip of the upper jaw, many along
the base of the dorsal-fin spines (usually including
spine 8, 9, or 10), and a well-speckled ventral
abdominal wall. These are typically all present
along with a minimal head complement of the simple
U or O. In those other species, one or more of the
combination is typically missing, especially when
the head pattern is limited to a U or O. In addition,
L. haitiensis larvae tend to have a wide-U
or O with the middle pair of melanophores more widely-spaced
than the rear pair, vs. a narrowing-U or O in the
other Labrisomus. Morphology is generally
required for a definitive separation: L. haitiensis
larvae consistently have long pelvic fins, more
than two-thirds the distance to the vent, with a
short third ray (always less than half the next
ray). Separation from the 18-spined species is easier:
L.
nuchipinnis, L.
conditus, and L.
cricota larvae apparently never have the
pair of spots behind the tip of the upper jaw or
more than two or three abdominal speckles (usually
they have none visible), and consistently have melanophores
on the inner cleithrum in the gill cavity as well
as outlining the lower caudal-fin segmented rays
(the latter rare on L. haitiensis larvae).
In addition, those three species also have a narrowing-U,
or often even a V-pattern of head melanophores (not
seen on L. haitiensis larvae), and a different
arrangement of spinous dorsal-fin base melanophores
(8,11-12,14+). L.
albigenys and L.
nigricinctus larvae are very lightly-marked,
missing most of the melanophores characteristic
of L. haitiensis.
Among the Malacoctenus, only M.
triangulatus can occasionally have larvae
with melanophores along the base of the spinous
dorsal fin and they share the long pelvic fins with
a short third ray and even sometimes overlap in
fin-ray counts with L. haitiensis. They can
also have the basic head pattern of two crescents
of three large melanophores per side but have additional
melanophores over all fore- and mid-brain quadrants
varying from tiny to large, usually ten per side
or more. The "peppered head" variant of
L. haitiensis larvae can resemble that pattern,
but have most or all of the additional melanophores
limited to the midbrain lobes. M.
triangulatus can be distinguished further
by the lack of ventral abdominal speckling and the
absence of the pair of melanophores behind the tip
of the premaxilla, located adjacent to the base
of the ascending process, that is characteristic
of some Labrisomus; note that the rare M.
triangulatus variant (likely in transition)
can have a cluster of small melanophores in that
area and a pair on the ethmoids a very short distance
away. In that case, the dorsal-fin outline (with
a notably short third-to-last spine) and long cirri
separate M.
triangulatus. Lastly, the row of internal
melanophores overlying the vertebral column is spaced
at one for each vertebra in M.
triangulatus vs. every third vertebra in
Labrisomus.
Transitional L. haitiensis larvae
are distinguished by fin-ray counts, persistent
larval melanophores (although these disappear rapidly
in this species), and the distinct pattern of metamorphic
melanophores on the body. The pair of melanophores
behind the tip of the upper jaw is shared only by
the 19- and 20-spined species, L.
gobio, L.
guppyi, L.
kalisherae, and L.
bucciferus, all of which develop a uniform
speckling of metamorphic melanophores on the body.
The transitional larvae of L.
nuchipinnis, L.
conditus, and L.
cricota develop connected reticulated lines
over the body instead of the isolated loops and
rings on L. haitiensis. Transitional
M.
triangulatus larvae have a generally similar
appearance, but their melanophore pattern is inverted
triangles, widest at the base of the dorsal-fin
elements. Transitional M.
versicolor larvae have a very similar transitional
pattern of isolated bars and shapes, but have long
third pelvic-fin rays (about 3/4 the length of the
longest ray), no pair of spots at the tip of the
jaw, notably long cirri, and very different fin
ray counts.
The juveniles of the 19- and 20-spined Labrisomus
are blunt-headed, compared to the 18-spined Labrisomus
(other than L.
albigenys), i.e. the snout length (bony
orbital margin to the tip of the closed jaw) is
less than two-thirds the bony orbit diameter. Small
juveniles of the blunt-headed Labrisomus
are difficult to identify to species, especially
since some species described with opercular ocelli
can have only a diffuse dark spot on the operculum
when young. In addition, those species described
without ocelli can often have an outlined dark spot
on the operculum (but not a thin-rimmed ocellus)
as juveniles.
Juvenile L. haitiensis have long pelvic-fins
with a short third ray that separate them from all
congeners (except L.
filamentosus) and also have high fin-ray
counts that rarely overlap with other Labrisomus
species. Juvenile L.
filamentosus have not been identified, but,
since juvenile L. haitiensis can also have
an ocellus-like spot on the operculum, it is possible
that L.
filamentosus may only be separated by fin-ray
counts: an extra dorsal soft-fin ray (dorsal elements
of 33) and 13 pectoral-fin rays (vs. 14). The marking
differences for juvenile L. haitiensis are
subtle, with L.
bucciferus and L.
kalisherae sharing the relatively long first
dorsal-fin spines (often with white-shaded membranes)
and the pigment on the caudal-fin base outlining
three light spots (vs. ending as a mostly straight
vertical line in L.
gobio and L.
guppyi). In addition, L.
bucciferus have fewer dark bands along the
pelvic fin-rays (4 vs. 6 or 7), L.
gobio differ in having the dark bars on
the body not extending onto the fins, and L.
guppyi rapidly develop a thin-rimmed opercular
ocellus and have a much shorter first dorsal-fin
spine (less than a third of the body depth at the
dorsal-fin origin vs. half or more). L.
albigenys differ by having a very short
first dorsal-fin spine and lower fin-ray counts.
The remaining Labrisomus juveniles are distinguished
by their longer snouts and lower fin-ray counts
i.e. L.
nigricinctus and L.
nuchipinnis (with well-delineated ocelli),
as well as L.
conditus and L.
cricota.
Labrisomus haitiensis
larva
18.0 mm SL, DNA-confirmed
ID
first dorsal spine 2.5
into body depth & similar length to mid-fin
spines
ventral abdominal speckling
Carrie Bow Cay, Belize,
7 Jan 1986
Labrisomus haitiensis
larvae
17.0-18.4 mm SL, DNA-confirmed
ID
range of head spot patterns
two are rare "peppered head" variants
Carrie Bow Cay, Belize,
7 Jan 1986
Labrisomus
haitiensis larva
19.0 mm SL
rare variant with caudal-ray
outlining, a characteristic of L.
nuchipinnis
Carrie Bow Cay, Belize,
7 Jan 1986
Labrisomus haitiensis
early trans. larva
17.0 mm SL, DNA-confirmed
ID
two long pelvic-fin rays
Magens Bay, St. Thomas
USVI ST509
Labrisomus haitiensis
transitional larva
16.9 mm SL, DNA-confirmed
ID
St. Thomas, USVI/J. Lamkin
& A. Shiroza
Labrisomus
haitiensis juvenile
21.1 mm SL, DNA-confirmed
ID
opercular pseudo-ocellus
Barbados, coll. by Henri
Valles
Labrisomus haitiensis
juvenile
29.2 mm SL, DNA-confirmed
ID
bars onto fins, three
pale spots at tail
first dorsal spine twice into body depth &
similar length to mid-fin spines
two long pelvic-fin rays, short third
Colon, Panama N7530a
Labrisomus filamentosus
Diagnosis: The modal
fin-ray count of D-XXI,12 A-II,21-22 and P-13 indicates
Labrisomus filamentosus. Only M.
boehlkei share the high dorsal-fin count,
but they have 15 pectoral-fin rays.
Ecology:
The quillfin blenny is an uncommon blenny found
on deep reef dropoffs. The species is found in the
Bahamas and most of the Caribbean, but not Florida,
the Gulf of Mexico, or the SW Caribbean across to
Venezuela. Unfortunately, the 19, 20, and 21-spined
Labrisomus are very similar morphologically
and are frequently confused in guidebooks and photographs.
All six species share the blunt-headed and goggle-eyed
appearance, at least as juveniles, i.e. the snout
length (bony orbit to jaw tip) is less than two-thirds
of the orbit diameter. Adult quillfin blennies are
easily recognized in the field by greatly-extended
spines at the front of the dorsal fin and a prominent
opercular ocellus. Juveniles may be difficult to
identify, since the distinguishing markings for
each species are either not well-developed or shared
by other species at this early stage. L. filamentosus
larvae are unknown in collections.
Description: Larval
L. filamentosus have not been identified.
Analogues:
Juvenile L. filamentosus have not been identified,
but, since juvenile L.
haitiensis share the long pelvic fins and
most markings and can also have an ocellus-like
spot on the operculum, it is possible that L.
filamentosus will only be distinguished by fin-ray
counts: their extra dorsal soft-fin ray (dorsal
elements of 33) and only 13 pectoral-fin rays (vs.
14).
Labrisomus filamentosus
Jamaica, Discovery Bay
Photo courtesy Mark Steele
Starksia
This genus of tiny and elusive blennies may become
one of the largest genera of Caribbean reef fishes
as more species are described in the coming years
(presently 25 and counting). Genetic analyses show
that many long-standing widespread species are made
up of complexes of similar-appearing species, usually
geographically separated (easy to explain), but
sometimes sympatric (not so easy to explain). Our
recent
paper describes seven new species, and numerous
other unnamed lineages.
Unfortunately, fin-ray counts for the entire genus
are similar and broadly overlapping. Furthermore,
adult markings on Starksia are both highly
variable and patterns are often shared among species,
making identifications particularly challenging.
It certainly does not help that males and females
often have different sets of markings. Much of the
variation in appearance occurs in live coloration
only and the dearth of live specimens or underwater
photographs makes identifications quite difficult.
Starksia are morphologically similar to
other labrisomids and some chaenopsid
blennies and are distinguished primarily by having
fewer soft dorsal-fin rays: 7 or 8, uncommonly 9.
Mature males have unusual genital papillae whose
morphology can be useful for taxonomy. Starksia
fin-ray counts can overlap the lowest range for
several species of Malacoctenus,
although only a very rare individual shares the
full combination of median and pectoral-fin ray
counts. Two common Malacoctenus
species occasionally have as few as 8 dorsal-fin
soft rays: M.
macropus (with 15 pectoral-fin rays) and
M.
erdmani (with 16). In general, the overlapping
Malacoctenus
species have higher pectoral and anal-fin ray counts.
Additional characters distinguishing Starksia
include fewer procurrent caudal-fin rays (4-5, vs.
6 or more), the last dorsal-fin spine stout and
long (2/3 to 3/4 of the first soft ray), and the
shortest dorsal-fin spine typically the third or
fourth-to-last.
Some of the larger-settling Starksia species,
particularly the S. ocellata complex, can
resemble M.
macropus as juveniles. They share long single
nuchal and orbital cirri and most fin-ray counts,
but Malacoctenus
have three pelvic-fin rays vs. two in Starksia.
M.
macropus also develop smaller mouths, pointier
snouts, and a narrower caudal peduncle (the maxilla
extends only to the anterior half of the orbit and
the caudal peduncle depth is less than half the
maximum body depth). Another useful distinction
is that Starksia have separate anterior and
posterior segments of the lateral line while other
labrisomids have continuous lateral lines. Juveniles
and adults of the other species of Malacoctenus
and Labrisomus
are also quickly excluded by multifid cirri and
larger size. Some chaenopsid
blennies are also small and can resemble Starksia,
however all have more than 10 dorsal-fin soft rays,
typically several more, and more total dorsal-fin
elements, 30 or more, usually many more (only Coralliozetus
cardonae would overlap the total dorsal-fin
count, but still ten or more soft rays). In addition,
chaenopsids
characteristically have the last dorsal-fin spine
the shortest and slender, while Starksia
have a distinctly long and stout last spine.
Most information is from Bohlke and Springer's
1961 monograph "A review of the Atlantic species
of the Clinid fish genus Starksia",
Greenfield's "..Starksia ocellata-complex.."
(1979), Greenfield & Johnson's 1985 large survey
paper on "The blennioid fishes of Belize and
Honduras...", Jeff Williams' 2003 review "Descriptions
of six new Caribbean fish species in the genus
Starksia (Labrisomidae)" by Williams and
Mounts (note that there are many post-publication
corrections by Jeff), and our recent 2011
review paper.
Starksia species list
(note: fin-ray counts in parentheses are ranges,
mode in bold, and two anal-fin spines not listed)
with 14 pectoral-fin rays:
The S.
atlantica complex, no orbital cirrus (all
other Starksia have an orbital cirrus) (shallow
forereef)
S.
atlantica in the Bahamas and Antilles
(18-19,7-8,15-16,14) and other locations
and lineages
S.
williamsi in the northern Lesser Antilles
(USVI and Saba)(19-20,7-8,16-17,13)
The S.
sluiteri complex (two habitat types): (shallow
coral, offshore reefs, continental and insular)
S.
sluiteri in the Neth. Antilles (19-20,7-8-9,15-16,13)
and other locations and lineages
S.
greenfieldi in the SE Caribbean (18-19-20,7-8-9,15-16,13)
S.
langi in the Bay of Honduras and Panama
(19-20,7-8,15-16,13)
S. multilepis in Noronha, Brazil (20,8,16-17,13)
(shallow rock, exposed, low island)
S. fasciata in the Bahamas and Cuba
(19-20,7-8,15-16,13)
S. y-lineata Cayman Islands & Nicaraguan
shelf (18-19-20,7-8,15-16,13)
S. smithvanizi in Navassa, USVI (19,7-8,15-16,13)
And others:
S. leucovitta in Navassa (18-19,8-9,15-16-17,13)
(deep rocky wall)
S. melasma in PR & USVI (18-19-20,7-8-9,15-16,13)
S. rava in Tobago (19-20-21,7-8-9,16-17-18,13)
(deep rocky wall)
S. sella in Tobago (18-19,7-8-9,16-17,13)
(deep rocky wall)
S. starcki is a distinctive species from
the Florida Keys, Yucatan & Belize (20(BZ)-21(FL),
8-9, 18(BZ)-19(FL),13)
Nemaclinus atelestos, a rare labrisomid
found only on deep dropoffs, looks much like the
Starksia species. It has higher median-fin
ray counts, but fewer pectoral-fin rays: D-XXI-XXIII,7-9
A-II,18-19 and only 11-12 pectoral-fin rays. It
also has notably long pelvic-fin rays, extending
well beyond the origin of the anal fin.
How to recognize
Starksia larvae
Larvae of Starksia are small typical-looking
labrisomids and have few distinguishing characteristics,
making them difficult to detect among the far more abundant
Malacoctenus
and Labrisomus
larvae in most samples. Fortunately, most Starksia
larvae are missing some of the typical labrisomid melanophores
and the exceptions have distinctive larval markings, such
as the irregular (larger and deeper) melanophores in the
anal row on larvae of the S.
atlantica and S.
ocellata complexes. In general, especially at earlier-stages,
Starksia larvae have more of a tadpole-like shape
with wider heads, than the other slimmer labrisomids and
the related tripterygiids
and chaenopsids
(note wide triangular forebrain lobes in Starksia
larva (right, larva on the left).
There are no taxognomonic characters
uniquely identifying all Starksia larvae, but
the absence of a set of common markings is typical
of most Starksia larvae. Typically, the only
anterior larval melanophore is at the isthmus. Notably,
most larvae are missing the cranial, deep nuchal,
cheek, internal otic, and pelvic melanophores (there
are exceptions for all but the cheek melanophore among
the S.
atlantica and S.
ocellata complexes). On all species, the anal
row spares one or more of the last rays and usually
there are no ventral caudal-peduncle melanophores
(at most one on some individuals of some species).
The absence of some of the usually consistent melanophores
on a labrisomid larva should prompt a closer examination. Chaenopsidae
larvae share the light marking patterns but have more
total pvm (19 or more) and more dorsal and anal-fin
soft rays.
Paraclinus
larvae superficially look much like Starksia larvae,
but can be distinguished by their distinctive all or all-but-one
spinous dorsal fin. They also have no preopercular spines
and more melanophores, including the deep nuchal and deep
pelvic and often the cheek and otic melanophores, as well
as a regular anal row with more than 18 total pvm. The Starksia
species that share deep nuchal (rarely) and pelvic melanophores
can be distinguished from Paraclinus
by having diagnostic irregular large or deep anal-row melanophores
and shorter anal rows). Starksia typically have 5
or 6 procurrent caudal-fin rays, one or two more than Paraclinus
larvae, and one to several fewer than the Malacoctenus
and Labrisomus.
Starksia with 14 pectoral-fin rays
Starksia atlantica complex
S. atlantica
S. springeri
S. sangreyae
Diagnosis: The modal
fin-ray count of D-XVIII-IXX,8 A-II,15-16 and P-14
indicates species of the Starksia atlantica
complex. S. elongata and the S.
ocellata complex share the 14 pectoral-fin
rays but have more dorsal-fin spines and anal-fin
rays. (DNA)
S. atlantica occurs in the Bahamas. S.
springeri is found in the Netherlands Antilles
and S. sangreyae is the species in the Bay
of Honduras. Other deeply-divergent DNA lineages
have been identified elsewhere in the Caribbean.
Description: Body long, narrow, and thin
with a large round eye, pointed snout, and medium
terminal mouth. Long continuous dorsal and anal
fins with a short and narrow caudal peduncle. Pectoral
fins long, reaching past the vent, and pelvic fins
long and thread-like, reaching about half-way to
the vent. There are three spines along the angle
of the preopercle that wane during transition. There
are often no cranial, deep nuchal, otic, or cheek
melanophores, but some larvae have cranial melanophores,
either a pair side-by-side or one of the pair, and
a few larvae have a mostly obscured deep nuchal
melanophore. There are no melanophores along the
dorsal-fin or caudal-fin base. There is an isthmus
melanophore as well as a surface pelvic melanophore
on most larvae: rarely the isthmus melanophore is
absent. The anal row is irregular and short: starting
behind the second spine and typically missing the
melanophores at the 3rd and 5th soft rays with a
larger, deeper and dendritic melanophore near the
base of the 4th ray. The row ends around the 11th
soft ray and is followed by another larger and deeper
melanophore near the base of the 13th soft ray.
There are no ventral caudal-peduncle melanophores
and only 9-12 total posterior ventral-midline melanophores.
Internal retroperitoneal melanophores are only visible
on less-developed larvae.
Transitional larvae develop metamorphic melanophores
in a pattern of bands over the head and a single
nuchal band, followed by a short bar below the eye
at 5 o'clock and a broad oblique stripe from the
eye across the anterior half of the jaws.
Analogues: Larvae
are distinguished from other labrisomid genera by
a wider interorbital region and a short anal row,
with only 9 to 12 total pvm (including irregular
deep melanophores), along with the absence of cheek
and otic and frequently the cranial and deep nuchal
melanophores (and the fin-ray counts detailed above).
Chaenopsid
larvae share the light marking patterns but have
more total pvm (19 or more) and more dorsal and
anal-fin soft rays. Larvae of the S.
ocellata complex have 14 to 18 pvm with
5 or 6 before the first deep melanophore, and higher
dorsal and anal-fin element counts. Larvae of the
S.
lepicoelia complex have 13 to 17 pvm, all
regular (some do have the last melanophore located
deep), are missing the pelvic melanophore, and have
13 pectoral-fin rays. Metamorphic melanophore patterns
are quite similar in the genus, but in larvae of
the S.
lepicoelia complex the patch of melanophores
over the head does not develop into well-defined
discrete bands and they develop prominent single
melanophores on the body on each side under the
gill covers.
Starksia atlantica
transitional larva
9.8 mm SL
Barbados, coll. by Henri
Valles
Starksia ocellata complex
S. ocellata
S. occidentalis
S. culebrae
S. guttata
S. variabilis
Diagnosis: The modal
fin-ray count of D-XX-XXI,8 A-II,17-18 and P-14
indicates species of the Starksiaocellata
complex. S. elongata can share this modal
count (modally 17 anal-fin soft rays) but usually
have fewer than 5 procurrent caudal-fin rays and
are more slender. The S.
atlantica complex also have 14 pectoral-fin
rays but have fewer dorsal-fin spines and anal-fin
rays. (DNA)
S. ocellata occurs in Florida and the Gulf
of Mexico. S. occidentalis is found along
the Central American coastline. S. culebrae
is the species in the N. Lesser Antilles, S.
guttata in the S. Lesser Antilles, and S.
variabilis in Colombia.
Description: (based on S. occidentalis)
Body long, narrow, and thin with a large round eye,
pointed snout, and medium terminal mouth. Long continuous
dorsal and anal fins with a short and narrow caudal
peduncle. Pectoral fins long, reaching past the
vent, and pelvic fins long and thread-like, reaching
about half-way to the vent. There are three spines
along the angle of the preopercle that wane during
transition. There are no cranial, deep nuchal, otic,
or cheek melanophores (but one transitional S.
ocellata larva has deep nuchal and otic melanophores).
There are no melanophores along the dorsal-fin or
caudal-fin base. There is an isthmus melanophore
as well as a pelvic melanophore on most larvae:
the pelvic melanophore is either deep (and sometimes
inconspicuous) or near the surface, not far from
the isthmus melanophore. A few have only the isthmus
melanophore, and rarely there is a surface pelvic
melanophore with no isthmus melanophore. A few larvae
have an additional tiny melanophore on the anterior
midline of the abdominal wall. The anal row usually
starts at the second spine and the series is irregular:
typically the melanophore at the 5th or 6th soft
ray is more prominent than the others (not deeper)
and the next one is often missing (on more than
half the larvae); on occasion, the one before the
larger one is missing, and uncommonly the row is
regular until the last deep melanophore. The row
usually stops at the 14th soft ray followed by a
deep larger melanophore underlying the base of the
last two rays. Uncommonly, there is an additional
small melanophore after the last ray on the caudal
peduncle. There are 14-18 total posterior ventral-midline
melanophores. Internal retroperitoneal melanophores
are only visible on less-developed larvae.
Transitional larvae develop metamorphic melanophores
in discrete bands across the top of the head, over
the fore and midbrain lobes, along with a nuchal
band, followed by a short bar below the eye at 5
o'clock and an oblique broad stripe from the eye
across the anterior third of the jaws. The cranial
metamorphic melanophores vary from three bands that
can split to six bands to a solid patch. Settled
juveniles have a dense and solid patch of melanophores
over the top of the head.
Analogues: Larvae
are distinguished from other labrisomid genera by
a wider interorbital region and the absence of cranial,
deep nuchal, cheek, and otic melanophores (rare
exceptions), as well as an irregular anal row, relatively
short with only 14 to 18 total pvm (and the fin-ray
counts detailed above). Chaenopsid
larvae share the light marking patterns but have
more total pvm (19 or more) and more dorsal and
anal-fin soft rays. Larvae of the S.
atlantica complex have a different pattern
of irregular anal-row melanophores with only two
or three before the first deep melanophore and only
9 to 12 pvm, and have fewer dorsal and anal-fin
elements. Larvae of the S.
lepicoelia complex can appear quite similar,
but are missing the pelvic melanophore, have no
irregular melanophores in the anal row (some do
have the last melanophore located deep), have 13
pectoral-fin rays (and lower dorsal and anal fin-ray
counts than the co-occurring member of the S.
ocellata complex). Metamorphic melanophore patterns
are quite similar, but in larvae of the S.
lepicoelia complex the patch of melanophores
over the head develops into poorly defined bands,
and they develop prominent single melanophores on
the body on each side under the gill covers.
Starksia occidentalis
larva
9.9 mm SL
San Blas, Panama, SB82-52
Starksia occidentalis
larvae
9.0-9.4 mm SL
irregular anal row melanophores
San Blas, Panama, SB86-405
Starksia occidentalis
larva
9.0 mm SL
note transverse septum
melanophore
San Blas, Panama, SB86-405
Starksia occidentalis
larvae
9.0 mm SL
isthmus, surface and
sub-surface pelvic
melanophores, note preopercular
spines
San Blas, Panama, SB82-082
Starksia occidentalis
larva
9.1 mm SL
isthmus, deep pelvic,
post-pelvic
melanophores, note preopercular
spines
San Blas, Panama, SB83-100
Starksia occidentalis
larva
9.0 mm SL
deep last mel and 5 procurrent
caudal
San Blas, Panama, SB82-082
Starksia occidentalis
larva
9.5 mm SL
isthmus and deep pelvic
melanophores
deep last melanophore
San Blas, Panama, SB82-082
Starksia occidentalis
transitional larvae
9.0-10.0 mm SL
head marking variation
San Blas, Panama, SB83-SB86
Starksia occidentalis
transitional larva
9.6 mm SL
head marking variation
San Blas, Panama, SB86-1008
Starksia occidentalis
transitional larva
10.0 mm SL
San Blas, Panama, SB86-608
Starksia ocellata
transitional larva
8.6 mm SL, DNA-confirmed
ID
deep nuchal melanophore
at transition
SE Florida, coll. Lance
Jordan
Starksia occidentalis
transitional juv
9.9 mm SL, DNA-confirmed
ID
Colon, Panama, May 29,
2007
Starksia with 13 pectoral-fin rays
Starksia lepicoelia complex
S. lepicoelia
S. robertsoni
S. weigti
S. williamsi
Diagnosis: The modal
fin-ray count of D-IXX-XX,7 or 8 A-II,16-17 and
P-13 broadly overlaps almost all of the numerous
13 pectoral-fin-rayed Starksia species. A
strong mode of D-XX,7 is (thus far) found only in
S. robertsoni, S. aff. lepicoelia
from Barbados, and S.
nanodes. Precise knowledge of the locality
and the sympatric congeners (or DNA sequencing)
is required for species-level identification among
this large group of species. (DNA)
S. lepicoelia was described from the Bahamas,
where more than one genetic lineage occurs. S.
robertsoni is found in Panama, S. weigti
is the species in the Bay of Honduras, and S.
williamsi occurs in the N. Lesser Antilles.
Other deeply-divergent DNA lineages have been identified
elsewhere in the Caribbean.
Description: Body long, narrow, and thin
with a large round eye, pointed snout, and medium
terminal mouth. Long continuous dorsal and anal
fins with a short and narrow caudal peduncle. Pectoral
fins long, reaching past the vent, and pelvic fins
long and thread-like, reaching about half-way to
the vent. There are three spines along the angle
of the preopercle that wane during transition. There
are no cranial, deep nuchal, otic, or cheek melanophores.
There are no melanophores along the dorsal-fin or
caudal-fin base. There is a single mid-isthmus melanophore
and no pelvic melanophores. The anal row starts
at the second spine or first soft ray and spares
one or a few of the next-to-last rays (sometimes
one or two others can be missing). The last melanophore
can be either just after the base of the last ray
or sometimes deep under the last two rays (Barbados).
There are 13-17 total posterior ventral-midline
melanophores. Internal retroperitoneal melanophores
are only visible on less-developed larvae.
Transitional larvae develop metamorphic melanophores
in a broad, mostly uniform patch over the head and
a single nuchal band, followed by a short bar below
the eye at 5 o'clock and a broad oblique stripe
from the eye across the anterior half of the jaws.
Transitional larvae develop one or two large surface
melanophores (that look more like larval than metamorphic
melanophores) on each side on the thorax under the
gill cover, below the pectoral insertion. As transition
progresses, the orbital cirrus grows and the fish
become mostly pallid (in life they are red), retaining
few dark markings. The prominent dense patch of
metamorphic melanophores over the head separates
into poorly-separated bands and then disperse (some
juveniles (Barbados) then develop a prominent midline
cranial melanophore between the forebrain and midbrain).
Analogues: Larvae
are distinguished from other labrisomid genera by
a wider interorbital region and the absence of cranial,
deep nuchal, cheek, otic, and pelvic melanophores,
as well as a relatively short row of 13 to 17 pvm
(and the fin-ray counts detailed above). Chaenopsid
larvae share the light marking patterns but have
more total pvm (19 or more) and more dorsal and
anal-fin soft rays. Larvae of the S.
ocellata complex have a similar number of
ventral melanophores and can be difficult to distinguish,
but usually have a pelvic melanophore, a more prominent
melanophore in the mid anal row with the adjacent
one often missing, and 14 pectoral-fin rays (and
higher dorsal and anal fin-ray counts than the co-occurring
member of the S. lepicoelia complex). S.
atlantica larvae are distinguished by only
9-12 pvm, a pelvic melanophore, sometimes cranial
melanophores, irregular deep melanophores in the
anal row, and 14 pectoral-fin rays. Metamorphic
melanophore patterns are also similar, but in the
S.
ocellata and S.
atlantica complexes the melanophores over
the head become well-defined discrete bands (before
coalescing into a dense patch in juveniles) and
they do not have the large thoracic melanophores
on the body on each side under the gill covers.
In addition, transitional larvae of the S.
atlantica complex do not develop an orbital
cirrus.
Starksia williamsi
early-stage larva
6.5 mm SL, DNA-confirmed
ID
St. Thomas USVI offshore
tow Aki
Starksia williamsi
early larva (left)
vs. Labrisomus gobio
(center)
vs. Enneanectes sp.
(right)
note no deep nuchal or
cranial mels
triangular
vs. oval forebrain lobes
DNA-confirmed IDs
St. Thomas USVI offshore
tow Aki
Starksia
robertsoni larva
8.2 mm SL
D-XX,7 A-II,16 P-13
San Blas, Panama SB83-164
Starksia aff.
lepicoelia transitional larva
8.2 mm SL, DNA-confirmed
ID
D-XX,7 A-II,16 P-13
Barbados, coll. Henri
Valles
Starksia aff.
lepicoelia transitional series
8.2. 9.8, 10.3 mm SL,
DNA-confirmed ID
Barbados, coll. Henri
Valles
Starksia aff.
lepicoelia transitional larvae
10.2, 9.8 mm SL, DNA-confirmed
ID
deep melanophore at end of anal row
Barbados, coll. Henri
Valles
Starksia aff.
lepicoelia transitional larva
10.2 mm SL, DNA-confirmed
ID
isthmus and thoracic
wall melanophore
Barbados, coll. Henri
Valles
Starksia weigti
transitional larva
9.8 mm SL, DNA-confirmed
ID
D-XIX,8 A-II,16 P-13
regular anal row, no
pelvic melanophore
thoracic wall melanophores
small spine on preopercular
margin
Turneffe, Belize, coll.
Cormac Nolan
Starksia nanodes
Diagnosis: The modal
fin-ray count of D-XX,7 A-II,16-17 and P-13 indicates
Starksia nanodes, as well as S.
robertsoni from Panama and S.
aff. lepicoelia from Barbados which share
the strong mode of D-XX,7. Precise knowledge of
the locality and the sympatric congeners (or DNA
sequencing) is required for species-level identification
among this group of species. (DNA)
Description: Body long, narrow, and thin
with a large round eye, pointed snout, and medium
terminal mouth. Long continuous dorsal and anal
fins with a short and narrow caudal peduncle. Pectoral
fins long, reaching past the vent, and pelvic fins
long and thread-like, reaching about half-way to
the vent. There are no cranial, deep nuchal, otic,
or cheek melanophores. There are no melanophores
along the dorsal-fin or caudal-fin base. There is
a single large isthmus melanophore and no pelvic
melanophores. The anal row starts at the fifth soft
ray and ends before the last several rays. Internal
retroperitoneal melanophores are only visible on
less-developed larvae.
Transitional larvae develop metamorphic melanophores
in a loose banded pattern over the cranium and a
short bar below the eye at 6 o'clock.
Analogues: Larvae
are distinguished from other labrisomid genera by
the absence of cranial, deep nuchal, cheek, otic,
and pelvic melanophores, as well as a wider interorbital
region and a relatively short anal row (and the
fin-ray counts detailed above). Larvae of the S.
lepicoelia, S.
ocellata, and S.
atlantica complexes are larger at transition
and the anal row extends to the end of the fin;
the latter two complexes have cranial and/or pelvic
melanophores and 14 pectoral-fin rays and variously
higher dorsal and anal fin-ray counts.
Starksia nanodes
transitional recruit
10.0 mm SL
Barbados, Henri Valle
2005R21
Starksia sluiteri complex
S. sluiteri
S. fasciata
S. langi
S. greenfieldi
S. y-lineata
S. smithvanizi
Diagnosis: The modal
fin-ray count of D-XIX,8 A-II,15-16 and P-13 broadly
overlaps almost all of the numerous 13 pectoral-fin-rayed
Starksia species. Precise knowledge of the
locality and the sympatric congeners (or DNA sequencing)
is required for species-level identification among
this large group of species. (DNA)
S. sluiteri is found in the Netherlands
Antilles. S. fasciata occurs in the Bahamas
and Cuba, S. langi is the species in the
Bay of Honduras and perhaps Panama, and S. greenfieldi
occurs in the SE Caribbean. The endemic species
in the Cayman Islands is S. y-lineata and
S. smithvanizi is found in Navassa and the
N. Lesser Antilles.
19,7 2,15 adult
Starksia sp. transitional
larva
7.8 mm SL
D-XIX,8 A-II,16 P-13
no melanophore at end
of anal row, very short first dorsal-fin spine
fin count not mode for
S. nanodes or S. sluiteri
San Blas, Panama, SB86-1008
Paraclinus
The genus Paraclinus has fewer species in
the western Atlantic than the three other labrisomid
genera discussed above (many more Paraclinus
species occur in the eastern Pacific). Juveniles
and adults are easily recognized by their dorsal
fin, which is either all spines or all-but-one spines
(with a single segmented ray at the end of the fin).
Combinations of fin-ray counts can identify most
species.
Three species are widespread and common in shallow
waters, but they are tiny and well-camouflaged and
thus rarely seen underwater: P.
nigripinnis (D-30+1 A-II,17-18 Pect-13),
P.
fasciatus (D-29-30 A-II,18-20 Pect-13),
and P.
cingulatus (D-28 A-II,15-16 Pect-12).
Two species live only on deep reefs and dropoffs
and are quite rare: P. barbatus (D-28-29+1
A-II,19-20 Pect-13, chin barbel), and P. infrons
(D-26-28+1 A-II,18 Pect-12, no nuchal cirri).
The remaining three species have unusual distributions
and comprise P. marmoratus, commonly found
only in Florida and NE Venezuela, rare in the Bahamas
and Belize (D-28+1 A-II,19-21 Pect-13, extended
first dorsal spines, often anal-fin ocellus; shallow
lagoon patch reefs); P. grandicornis, found
only in Florida, the Bahamas, and the Virgin Islands,
living within sponges (D-27+1 A-II,18 Pect-12, large
orbital cirrus); and P. naeorhegmis from
the Bahamas, rare in Belize and Providencia (D-26-27
A-II,15-16 Pect-13; well-washed channels). Originally
described with very narrow ranges, more recent surveys
and photograph collections reveal that most species
(or, more likely, allopatric sets of cryptic species)
are wide-ranging throughout the region.
How to recognize
Paraclinus larvae
Pre-transitional larvae of Paraclinus
are relatively small and often resemble generic labrisomid
larvae, with no distinctive morphology or markings. The
one diagnostic feature is that the posterior dorsal fin
is made up of spines instead of soft rays. On well-developed
and well-preserved larvae this feature easily confirms the
identification, however the posterior spines of Paraclinus
tend to be broader than typical spines and are recognized
as spines mostly by the absence of segment divisions and
the bulb at the base of soft rays. On less-developed larvae
and larvae in poor condition, this distinction can be difficult
to assess.
An additional character quickly separating Paraclinus
larvae from the far more abundant larvae of Malacoctenus
and Labrisomus is the absence of cranial
melanophores. Paraclinus larvae also have
no obvious preopercular spines, while many immature
Malacoctenus
and Labrisomus
(and Starksia)
larvae have preopercular spines at the size of overlap
with Paraclinus larvae. A useful character,
even on poorly preserved larvae, is the number of
procurrent caudal-fin rays: Paraclinus larvae
have only 3 or 4 vs. 6 to 10 in Malacoctenus
and Labrisomus
(and 5 to 6 in Starksia).
Other than the diagnostic dorsal-fin
spine complement, Paraclinus larvae can be
distinguished from the similar Starksia
larvae by the consistent presence of the deep nuchal
and deep pelvic melanophores and often the cheek melanophore.
Deep melanophores, however, are not always visible
on well-developed larvae. Interestingly, transitional
Paraclinus larvae collected at night over the
reef do not have the metamorphic melanophore patterns
over the head typical for transitional Starksia
larvae and frequent on larvae of Malacoctenus
and Labrisomus.
Nevertheless, by the morning (in collections made
by unattended crest nets), metamorphic melanophores
have developed.
Paraclinus fasciatus
Diagnosis: The modal
fin-ray count of D-XXIX A-II,18-19 and P-13 (dorsal
fin all spines and only two pelvic-fin soft rays)
indicates Paraclinus fasciatus (almost all
have 29-30 spines A-II,18-19). Two other Caribbean
species have the dorsal fin all spinous (also only
two pelvic-fin rays): P.
cingulatus, with 12 pectoral-fin rays (28
spines, A-II,15-16), and P. naeorhegmis (26
spines, II,15-16) with 13 pectoral-fin rays. Paraclinus
larvae are common in my collections from Panama
and the vast majority are P. fasciatus. (DNA)
Description: Pre-transitional larvae: Body long,
narrow, and thin with a medium round eye, pointed
snout, and terminal medium mouth. Larvae can range
from hunched-over to long and straight. Long continuous
dorsal and anal fins with a very short and narrow
caudal peduncle. Pectoral fins medium, reaching
to the vent, and pelvic fins relatively long and
thread-like. In later larvae, a notch develops in
the membrane between the first three dorsal-fin
spines and the rest of the fin. There are no surface
melanophores along the dorsal aspect of the body
(a rare sample will have some larvae with melanophores
at the base of one or a few of the last dorsal-fin
spines). There is usually a cheek melanophore on
each side. There are no melanophores along the caudal-fin
base. Along the ventral midline there are melanophores
at the isthmus and deep behind the pelvic-fin base.
Along the anal fin there is a melanophore at the
base of each anal-fin soft ray (sometimes also at
the second spine and sometimes sparing the last
and/or second-to-last ray), closely followed by
one to three along the ventral midline of the caudal
peduncle (uncommonly none), with the last in the
series often on the rim of the procurrent caudal
finfold. A small fraction of larvae (0-20% of larger
samples) have additional markings: a small midline
melanophore on the abdominal promontory just forward
of the vent and/or another or two midway along the
abdomen and/or (rarely) a melanophore on the lower
inner cleithrum inside the gill cavity on each side.
Internal melanophores comprise only the basic complement:
the nuchal midline (often obscured), otic capsule,
and overlying the abdominal organs.
Transitional stage: P. fasciatus
larvae are not caught at night over the reef with
metamorphic melanophores, which apparently must
develop after settlement. Transitional larvae often
have developed cirri and thicker bodies without
any change in markings. Some larvae do have additional
melanophores along the ventral midline of the abdomen
and in the gill cavity, but they are not the most
developed larvae in the collections and these markings
are present on early-stage larvae as well. Rarely,
larvae can also have melanophores at the base of
a few spines in the last third of the dorsal fin,
or one or two under the eye, or one along the anterior
portion of the lower jaw; these may represent transitional
changes.
Analogues:
Larval P. fasciatus are not easily distinguished
from larvae of the other common species, P.
nigripinnis. Unfortunately their fin-ray
counts can overlap and the diagnostic differences
only become clear in well-developed larvae. P.
fasciatus larvae are distinguished by having
all spines in the dorsal fin (vs. a last soft ray),
a relatively long pelvic fin reaching more than
halfway to the vent with only two pelvic-fin soft
rays (vs. shorter and a small third ray), and usually
one or two more anal-fin rays. Unfortunately the
last dorsal-fin soft ray and the third pelvic-fin
ray can be difficult to discern without staining
the fish (and some P.
nigripinnis never form the soft ray) and
there is some overlap in the length of the pelvic
fins before transition. Furthermore, both species
have many individuals with 18 anal-fin soft rays.
It is likely the two species are only reliably distinguished
by eye when in good condition and near the settlement
stage.
Paraclinus fasciatus
larva
8.8 mm SL
San Blas, Panama, SB86-501
Paraclinus cingulatus
Diagnosis: The modal
fin-ray count of D-XXVIII A-II,15-16 and P-12 indicates
Paraclinus cingulatus. Two other Caribbean
species have all dorsal-fin elements spinous (all
also with only two pelvic-fin rays), but both with
13 pectoral-fin rays: P. fasciatus, almost
all with D-29-30 A-II,18-19, and P. naeorhegmis
with 26 spines and 15-16 anal-fin soft rays. (DNA)
Description:
Paraclinus fasciatus
larva
8.8 mm SL
San Blas, Panama, SB86-501
Paraclinus nigripinnis
Diagnosis: The modal
fin-ray count of D-XXX,1 A-II,17-18 and P-13 indicates
Paraclinus nigripinnis. Several species share
the single segmented ray as the last dorsal-fin
element and all of these also have a small third
pelvic-fin soft ray (vs. only two), although only
P. marmoratus (D-28+1, A-II,19-21) and P.
barbatus (D-28-29+1 A-II,19-20) share the mode
of 13 pectoral-fin rays. (DNA)
Description: Pre-transitional larvae: Body long,
narrow, and thin with a medium round eye, pointed
snout, and terminal medium mouth. Larvae can range
from hunched-over to long and straight. Long continuous
dorsal and anal fins with a very short and narrow
caudal peduncle. Pectoral fins medium, reaching
to the vent, and pelvic fins relatively long and
thread-like. In later larvae, a notch develops in
the membrane between the first three dorsal-fin
spines and the rest of the fin. There are no surface
melanophores along the dorsal aspect of the body.
There is usually a cheek melanophore on each side.
There are no melanophores along the caudal-fin base.
Along the ventral midline there are melanophores
at the isthmus and deep behind the pelvic-fin base.
Along the anal fin there is a melanophore at the
base of each anal-fin soft ray (sometimes also at
the second spine and sometimes sparing the last
and/or second-to-last ray), closely followed by
one to three along the ventral midline of the caudal
peduncle (uncommonly none), with the last in the
series often on the rim of the procurrent caudal
finfold. Some larvae have additional markings: a
small midline melanophore on the abdominal promontory
just forward of the vent and/or another or two midway
along the abdomen and/or (rarely) a melanophore
on the lower inner cleithrum inside the gill cavity
on each side. Internal melanophores comprise only
the basic complement: the nuchal midline (often
obscured), otic capsule, and overlying the abdominal
organs.
Transitional stage: P. nigripinnis
larvae are not caught with metamorphic melanophores,
which must apparently develop after settlement.
Transitional larvae often have developed cirri without
any change in markings. Some larvae have additional
melanophores along the ventral midline of the abdomen,
but they are not the most developed larvae in the
collections and the abdominal markings are present
on early-stage larvae as well.
Analogues:
Larval P. nigripinnis are not easily distinguished
from larvae of the other common species, P.
fasciatus. Unfortunately their fin-ray counts
can overlap and the diagnostic differences only
become clear in well-developed larvae. P. nigripinnis
larvae are distinguished by having the last element
of the dorsal fin a soft ray (vs. all spines), a
relatively short pelvic fin reaching less than halfway
to the vent with a tiny third ray (innermost) (vs.
longer than halfway and only two soft rays), and
usually one or two fewer anal-fin rays. Unfortunately
the last dorsal-fin ray and the third pelvic-fin
ray can be difficult to discern without staining
the fish (and some P. nigripinnis never form
the last dorsal-fin ray) and there is some overlap
in the length of the pelvic fins before transition.
Furthermore, both species have many individuals
with 18 anal-fin soft rays. It is likely the two
species are only reliably distinguished by eye when
in good condition and near the settlement stage.
Paraclinus nigripinnis
larva
7.9 mm SL
San Blas, Panama, SB86-616
Paraclinus nigripinnis
larva
8.3 mm SL
pelvic-fin base melanophore
obscured
note last dorsal-fin element soft
and small third pelvic ray (innermost)
San Blas, Panama, SB86-507
Paraclinus nigripinnis
larva
8.3 mm SL
deep melanophore behind
brain
pelvic fin less than halfway to vent
small third pelvic-fin ray
no spines on preopercle